- SUGGESTED TOPICS
- The Magazine
- Newsletters
- Managing Yourself
- Managing Teams
- Work-life Balance
- The Big Idea
- Data & Visuals
- Reading Lists
- Case Selections
- HBR Learning
- Topic Feeds
- Account Settings
- Email Preferences

How to Give a Killer Presentation
- Chris Anderson

For more than 30 years, the TED conference series has presented enlightening talks that people enjoy watching. In this article, Anderson, TED’s curator, shares five keys to great presentations:
- Frame your story (figure out where to start and where to end).
- Plan your delivery (decide whether to memorize your speech word for word or develop bullet points and then rehearse it—over and over).
- Work on stage presence (but remember that your story matters more than how you stand or whether you’re visibly nervous).
- Plan the multimedia (whatever you do, don’t read from PowerPoint slides).
- Put it together (play to your strengths and be authentic).
According to Anderson, presentations rise or fall on the quality of the idea, the narrative, and the passion of the speaker. It’s about substance—not style. In fact, it’s fairly easy to “coach out” the problems in a talk, but there’s no way to “coach in” the basic story—the presenter has to have the raw material. So if your thinking is not there yet, he advises, decline that invitation to speak. Instead, keep working until you have an idea that’s worth sharing.
Lessons from TED
A little more than a year ago, on a trip to Nairobi, Kenya, some colleagues and I met a 12-year-old Masai boy named Richard Turere, who told us a fascinating story. His family raises livestock on the edge of a vast national park, and one of the biggest challenges is protecting the animals from lions—especially at night. Richard had noticed that placing lamps in a field didn’t deter lion attacks, but when he walked the field with a torch, the lions stayed away. From a young age, he’d been interested in electronics, teaching himself by, for example, taking apart his parents’ radio. He used that experience to devise a system of lights that would turn on and off in sequence—using solar panels, a car battery, and a motorcycle indicator box—and thereby create a sense of movement that he hoped would scare off the lions. He installed the lights, and the lions stopped attacking. Soon villages elsewhere in Kenya began installing Richard’s “lion lights.”
- CA Chris Anderson is the curator of TED.
Partner Center
9 Killer Speech Openers to Start a Talk or Presentation.

Danny Riley 8 min read
What you’ll learn:
- The importance of a “killer” speech opening.
- 9 powerful speech openers and how to use them.
- Examples from great speakers you can learn from.

Great speech openers hook your audience.
“ Well begun is half done” – Mary Poppins.
A killer speech opener will make the difference between a presentation that makes you soar or your audience snore .
I’ve researched the whole web to find nine killer speech openers to make your audience lean in and listen rather than tune out and daydream.
You’ll see how masters of the craft have used them, and how you can too.
Number seven takes hutzpah to pull off. Ready for the whole list of killer speech openers?
The Shock Opener
One of the best ways to open your speech with a buzz is to startle or shock them.
You can shock an audience in many ways, but they all rest on the major senses of VAKS:
- Kinesthetic (touch)
We don’t want your audience tasting your talk, but it should leave a good taste in their mouths.
Changing Minds suggests asking if the audience is awake after appearing from a flashbang and a cloud of smoke, and this might work for you if you’re a magician or playing some kind of character for your speech like a genie.
Suppose you aren’t going for the magic angle.
In that case, you can shock them on a psychological level instead, as Conor Neill recommends, and tell your audience a surprising fact or statistic that makes them question their thinking or beliefs.
“Did you know that half the water on earth is older than the sun?”
Questions like these will shake an audience awake and turn on their critical thinking nervous-system.
Don’t take my word for it; you can see an incredible demonstration of the shock opener in Mohammed Qahtani’s speech, The Power of Words .
Qahtani opens by taking out a cigarette and placing it into his mouth before trying to light it. The audience is so shocked that they gasp and tell him to stop.
Remember, if your audience is shocked, they are listening.
Your audience doesn’t always have to be jolted to attention with a shock opener, though you can use a more subtle approach to grab their focus.
Ready to speak with confidence ? Explore our training options...
The story opener.
You can set the tone of your speech instantly with a story .
In Hollywood, filmmakers and directors use an ‘establishing shot’ to set the tone and theme of the entire film.
When creating your speech, think of a short story that sums up your talk.
Maybe you tell half the story to begin with, and then the other half at the end.
The important thing is your tale must be relatable . If your audience can’t imagine themselves in the story, they won’t be engaged.
We all experience very similar things in life:
- We all went to school and had a teacher we loved
- We all have parents who loved us or made mistakes in our upbringing
- We all had a first crush.
We are all cut from the same cloth, so it’s good to be reminded that others are going through what we face or think as we do.
Bryan Stevenson does a stellar job of recounting his mischievous grandmother in his TED talk, We need to talk about an injustice .
The best thing is, you can combine a story-opener with any other speech opener in this list.
It’s truly versatile.
One of my favourite speech openers is next, though.
The Intrigue Opener
I love this speech opener.
What better way to hook your audience than to intrigue them with mystery or a juicy secret?
Take a look at Daniel Pink’s TED Talk The puzzle of motivation . After he begins, Pink, looking like a guilty man sent to the gallows tells his audience:
“I need to confess something, at the outset here. A little over 20 years ago, I did something I regret. Something I’m not particularly proud of”.
Wow. How intriguing, right?
You have to admit; you want to know what he’s about to confess.
Choose every sentence, every word, and every mark of punctuation to increase the tantalisation temperature.
Whether it’s a secret or confession, the Intrigue Opener piques just enough curiosity in your audience to keep them from checking Whatsapp.
As humans, we need closure.
We do not like open loops.
That’s why it is both enthralling and aggravating when someone plays on our need to be sure.
Just as we cannot stand an open loop, we are instantly engaged when someone gives us a puzzle to solve.
You’ll notice the best speeches, books, tv shows, and films do not spoon feed you all of the information.
I’ve always liked the way Malcolm Gladwell writes his non-fiction books because they contain puzzles that you solve as a reader.
This puzzle needs to be related to the speech or presentation you’re delivering, of course. It cannot be a random puzzle and will ideally be impossible or extremely difficult to solve at first.
After the speech begins and the puzzle is revealed, you should slowly drop hints on how to solve the mystery.
Up next, speech openers that use a physical object to create curiosity in the audience’s mind.
The Prop Opener
One of the most potent ways you will captivate your audience is to use a powerful prop in your opening address.
What better way to capture an audience’s imagination than to show them a mysterious or beautiful object?
If you’ve never seen the Prop Opener done well, then take a look at one of the greatest speeches of all time:
Dananjaya Hettiarachchi’s, See Something .
Danajaya enters with a simple rose in his breast pocket, takes it out, gazes at it nostalgically, smells it and then begins to speak.
This same prop appears again right at the end of his speech to end his talk with a flourish.
There are many different props you can use.
JJ Abrams used a Mystery Box to absorb the audience’s attention and used the box as a metaphor for his entire career.
If you think the prop opener is just for TED Talks and Toastmasters Final Speeches, remember that most company product launch centre around one or more props.
Steve Jobs revealed his new products in ever-innovative ways.
Still, while the last two speeches I’ve mentioned opened with physical items, most of Jobs’s presentations built intrigue through the sight of the product.
So remember, you can use an object, or tease your audience with the absence of a prop, but make that prop integral to your talk.
You don’t always have to use a prop, of course.
A more minimalist approach to opening your speech uses the best audience reaction a speaker can receive: laughter.
The Funny Opener
Using laughter to win over your audience is the golden ticket to immediate rapport with your audience.
Jack Schafer, PhD at Psychology Today, said that People Will Like You If You Make Them Laugh , which seems obvious, but at least you know we have scientists on the case.
He also mentions that constructing humour requires and projects a high level of intelligence .
Of course, laughter is subjective, but it is also infectious, and if you get enough members of your audience to titter, it will spread across the whole group.
If you want to see just how quickly you can win an audience over with humour, take a look at Ken Robinson’s subtle but delightful ability to raise a chuckle in his speech Do Schools Kill Creativity?
Ken’s ability to speak conversationally to an audience of thousands is genuinely remarkable.
If you break down his humour, it is easy to see how you could include similar content in your presentations.
Whether you can pull it off as well as Ken is another story.
Not everyone feels like they can be a comedian, though; I get that.
Well, that’s alright because there are other ways to open your talk that play on other strong emotions.
You can inspire your audience too.
The Inspirational Opener
One of my favourite ways to help beginner speakers to open their presentation is with a quote.
A quote acts like a story in that it sets the tone and theme of your speech, but it takes much less effort and even less skill.
An effective quote is usually only one line long and supported by the credibility of the original author who uttered those words.
Watch the way Clint Smith opens his TED Talk The Danger of Silence .
Using Martin Luther King’s voice to start his speech gives Clint what psychologists call the transference effect .
Just by citing someone else, especially someone admired and famous, you redirect the emotions an audience have towards that person onto yourself.
One caveat to using quotes, though:
Fact check them . I cringe whenever I see someone incorrectly quoting someone.
Have you ever heard the quote by Albert Einstein:
“Insanity is doing the same thing over and over, and expecting different results”?
A great quote, isn’t it?
But Albert Einstein never said those words .
A quick check on Reuters will help you add more credibility to your inspirational opener.
Finally, try to use a quote few people have ever heard. Inspiring words have been filling the archives of history for millennia, so seek out something that has been left dusty on the shelf rather than the same recycled iterations.
Next, let’s look at a type of bold speech opener that take real hutzpah to land well.
The Perspective Shift Opener
A powerful speech opener that will take confidence is the perspective shift opener.
This opener will lead the audience in one direction before changing direction and setting a new pace for the speech.
Cameron Russel does a fantastic job of controlling the frame in her TED Talk Looks aren’t everything. Believe me; I’m a model.
Russel takes to the stage dressed in a skimpy dress and begins to tell the audience about her career, but then does a rapid wardrobe change on stage in front of the entire audience.
This change of dress sets a new tone, feel, and direction for the speech.
If you can change the audience’s perspective or frame of reality, you are in the driving seat.
One of the best things you can hope for as a speaker is moving hearts and changing minds.
If you aren’t a confident speaker, start small.
Vanessa Van Edwards suggests never mentioning how nervous you are.
It’s distracting and makes the audience pick up on all the subtle nervous energy and cues you give off. Control the frame instead and act cool and confident: they will buy into it.
Another great way to hold frame control over an audience is by using the power of silence .
The Silence Opener
Silence is a valuable commodity in today’s noisy and distracting digital world.
Creating silence at the beginning of your talk can profoundly affect your audience and their focus.
Did you ever have a teacher at school who used silence effectively?
When my English classmates were noisy, our teacher Mr Rylance would hold up his hand in silence.
Slowly we would settle down and focus on his raised hand.
A few would giggle, but that would peter out until we all wrapt in a hypnotic stillness.
If you want to see an example of how to use silence, then look at Neal Glitterman’s speech The Power of Silence .
You can see how much gravity silence can have , especially as a speech opener.
The final killer opener I want to introduce you to is the big promise opener.
The Big Promise Opener
I believe that all speeches and presentations should contain a big promise as it tells your audience why they should keep on listening.
Ideally, your big promise will be your speech title or phrase that pays which is a recurring foundational phrase you will use throughout your presentation.
A big promise is your way of making a deal with the audience : you listen to me, and you’ll get something in return.
Creating a big promise at the beginning of your speech is like adding a teaser trailer to the beginning of a TV show. It suggests a reason you should stick around.
When Arthur Benjamin introduces his talk Faster than a calculator by announcing:
“I am a human calculator!”
You know that proof is on the way.
Remember the essential rule of the Big Promise Opener: make it big and keep your promise.
WOW your audience with these killer speech openers.
I hope you feel that I kept my promise of sharing nine killer speech openers to start a presentation.
Did you notice any other speech openers at the beginning of this article?
Don’t forget; these openers can be mixed and matched.
You can include a number of these speech openers in the same presentation to create more impact.
Let me know which of these killer openers was your favourite, and let me know if you have any more you’d like to share.
– Danny Riley
Join 350+ leaders getting my weekly tips on confidence and charisma... 👇
Navigation:.
Home About Success Stories Contact Privacy Policy
Work with Ed:
1-to-1 Coaching 1-Day Masterclass Team Training
Connect & Follow:
Get started:, copyright © 2023 project charisma ltd. all rights reserved..

How to Give a Killer Presentation [2024]
January 05, 2024
When you give a killer presentation, you stand a much greater chance of making it easy for your audience. And if you make it easy for your audience they are more likely to respond.
What does creating a killer presentation mean in reality.
By killer presentation, I mean:
- A presentation that can be remembered.
- A presentation that positions you as someone of note within your organisation.
- A presentation that creates action, change in behaviour or change in thinking
To create a killer presentation you need to be different, and you need to take a few risks. (If there was a simple formula, then everyone would be doing it.) But when I say take risks, I mean doing things that are done outside the typical business setting but are actually best practice presentation skills training shortcuts. Shortcuts that have been tried and tested for hundreds if not thousands of years. You want to follow the cardinal rule of ‘Don’t be boring’.
An Example of a Killer Presentation
John is the COO of a large utilities company. We trained him to improve his presentation skills. To deliver killer presentations. He has thousands of employees working daily in dangerous conditions, keeping water running for millions of customers. People have to work at heights, at depth, in the middle of the road, day and night, winter and summer. They work in every dangerous situation you could imagine.
And it is hard keeping everyone safe. Teams are scattered over several hundred square miles and they work autonomously, going to emergencies as and when they happen.
Health and safety is important to the business; it takes up a large part of senior management time. Every year the senior team gathers, and this year safety was again one of the main topics of the day.
John, was new as COO and determined to make an impact. He know that the presentation skills he learned in his training would be critical.
His HR team had pulled together a big presentation deck discussing health and safety. It had the company history, the key imperatives, the company values, best working practices and updates on recent legislation. It was a comprehensive pack and it had taken weeks to put together. They even brought in professional designers to make sure the presentation looked as important as it was.
John knew from his presentation skills training at Benjamin Ball Associates that starting his presentation with slides – no matter how good – would not work. So he used a story from his early career. He started his killer presentation like this:
“When I was in twenties I was put in charge of a building site. In my first week on that job we had a crane collapse. “That night, I had to knock on a woman’s door and tell her that her husband had been killed on a site where I was in charge. “I don’t want any of you to have to go through what I went through that day. That’s why we are talking about health and safety today.”
In less than 80 words John had grabbed his audience’s attention, had appealed to their emotions and made them realise that he knew what he was talking about.
That was a great example of a killer presentation using advanced presentation skills.
While not every presentation you do will be a killer presentation, it is something that you should aim for. Much better to aim high and fall short than aim low and fall short anyway.
Contact us now for a free consultation
Why is it hard to create killer presentations?
One of the big challenges we face in business is that The Presentation has become a standard way of communicating. Unfortunately, the average presenter and the average presentation is not very good. As a result, time is wasted, money is wasted and we are not as clear and inspiring as we should be.
The average business presentation is completely forgettable. I often challenge clients to count the number of presentations they have seen in the last year, and then to describe any memorable ones. I am lucky if they can speak about one.
Too many people, when they hear the word “presentation”, reach for their computer and start creating slides. They feel as if they are making progress.
They pull information together, they lay it out into a PowerPoint document and admire their handiwork. Then they start to think about how to tell the story. You’ll never give a killer presentation like this.
“Before anything else, preparation is the key to success.” – Alexander Graham Bell
What you need – A story, a script and a plan to create a killer presentation
BUT…the process for creating a great presentation should be more like that of making a movie. When you make a movie, filming (the equivalent of creating slides) only happens after you have a story, a script and a plan.
The better you have planned your presentation, the easier the process of giving it. At first, it may feel frustrating that you are not writing or creating slides. But when you invest in proper presentation planning, you will benefit many times over.
In the end you will spend less time playing with PowerPoint. You will spend less time editing and you will spend less time searching for a way to link the sections of your presentation together. You will probably find that practising and rehearsing is also easier and more enjoyable.
The other great advantage of improving your presentation skills is that you have formal check points where you can share your work to check that you are on track. This will help with timekeeping and, where necessary, make sure your colleagues are onside.
“There are three things that are important for a film. Number one is story, number two is story, number three is story. Good actors can save a bad script and make it bearable, but good actors can’t make a bad script good – they can just make it bearable.” – Mark Strickson, TV producer & acto r
Summary – how to improve your presentation skills for success
Start by planning, not by writing. This may feel counter intuitive, but you’ll make progress faster.
How do you give a killer presentation? Try these steps:
- Use AIM as the first stage of preparing your presentation
- A – Audience: Analyse your audience and understand their needs
- I – Intent: Be clear on the single purpose of your presentation.
- M – Message: Decide your one take-away message from your presentation
- Decide the three parts of your presentation
- Create and perfect a 90 second summary of your presentation before fleshing it out
- Answer the WHY questions in your presentation before the WHAT or HOW questions
- Imagine your audience asking So What? and What’s in it for me? throughout.
- Check your presentation summary against your Audience, Intent and Message.
Create a killer presentation. Step #1 – Have you taken AIM?
Summary – aim is an easy-to-apply planning tool that makes your business presentations and talks easier to prepare..
What typically goes wrong? Most people create presentations without proper planning. They start writing or creating visual aids before they have decided what they want to say.
Why does this matter? Without an effective tool for preparing a business presentation you waste time and will be less effective.
So, instead, Use AIM. Start with a blank sheet of paper and write the three letters AIM across the top. In each of these columns start writing what you know about A: Your Audience, I: Your intent, or Purpose and M: your take-away Message. For Audience ask yourself searching questions about why they are here and what they want from your presentation. For Intent, summarise your intent into one clear line. And for message identify the single message that you’d like your audience to take away from your business presentation. See the next sections for more detail on A, I and M.
“90% of how well the business presentation will go is determined before the speaker steps on the platform.” – Somers White
Improve your presentation skills tips
- Use the AIM approach before all communications.
- Keep working at AIM until you are happy.
Create a killer presentation. Step #2 – AIM Part 1: Who’s your audience and what do they need?
Summary – your audience, not you, should be centre of attention in your presentation. the better you understand your audience, the better your business presentation will be..
What typically goes wrong? When people give presentations about their latest project, they talk about their latest project. If they are reporting quarterly results, they report quarterly results. If they are speaking about their new business, they tell the audience about their new business. If explaining a new piece of regulation, they talk about elements of that regulation.
The problem with this approach is they are not including their audience in their presentation. And if they don’t include the audience, The audience will be less engaged.
Why does this matter? Audiences are selfish. They like being talked about.
Instead, to improve your presentation skills your presentation should be about what your subject means for your audience. For example:
- “What you can learn from our latest project.”
- “Our quarterly results and what they mean for your department next quarter.”
- “How our new business can make you money.”
- “What the new regulations mean for you and your clients”
To do this, you must understand your audience. That means asking questions about them and getting under their skin. For example, some questions you may have could include:
ABOUT BACKGROUND
- Who is coming to this business presentation?
- What common reference points can I use?
- What experiences have they shared?
ABOUT MOTIVATION
- Why are they coming?
- What problems do they have?
- What do they need and want?
- What will make life easy for them?
ABOUT EXPECTATIONS
- What would they like me to business presentation about?
- What would a win feel like for them?
- What will make them sit up?
ABOUT CONNECTING
- What can I say that will show them I am on their side?
- What stories will resonate?
- How can I add value?
ABOUT EMOTIONAL POSITION
- What frame of mind will they be in?
- What should I avoid talking about?
- What will make them feel good?
- What can I say at the start to win them over?
ABOUT MAKING IT EASY FOR THEM
- What specific language should I use?
- How should I position what I am talking about for this audience?
- What phrases will resonate?
The more you learn about your audience, the better you know them and the better you can plan your presentation for them.
“Designing a presentation without an audience in mind is like writing a love letter and addressing it: To Whom It May Concern.” – Ken Haemer, presentation designe r
Top Presentation Improvement Tips
- Before any presentation, analyse your audience
- Research them
- Make sure you really know them and their needs before you start planning what to say
Create a killer presentation. Step #3 – AIM Part 2: What are you trying to achieve?
Summary – decide early the intent or purpose to your presentation. this will help you direct your efforts and target your presentation so that you achieve your goals..
What typically goes wrong? “I’m going to talk about…” is a typical answer to the question “Why are you doing this business presentation?” But presenting about something is of no use to anyone. It is pointless.
For example:
- “I’m presenting about our new project”
- “I’m presenting about the new regulations”
- “A pitch about our new fund”
- “An introduction to ABCX co”
- “Monthly board report”
Why does this matter? For a presentation to work it requires a clear purpose. When you know your purpose you can harness your presentation to achieving just that.
So, instead, decide your intent. For example, when I asked a Chief Financial Officer recently what was the intent of his presentation, he was clear: he said that he “Wanted to look like the next CEO of this business.” This clear purpose made it easy to help him prepare what he said, how he said it and how he positioned himself.
Someone recently, when giving a presentation about new regulations, was clear that she wanted “to help companies use the new regulations to run better, more profitable businesses”.
An HR director who was introducing a new expense system was clear that her intent was to “get people to use the new system by next month so they can get paid faster and with less effort.’
A fund manager who was pitching a first time fund to new investors had a clear intent of “getting onto their radar screens and securing a second meeting.”
A company looking for a trade buyer had crystallised their intent into “creating excitement about the potential value of buying this business and demonstrate the risk of others buying it.”
“An accountant at an FMCG firm had the intent with his monthly board reports to “Get them to recognise the value my team adds.”
Having a clear intent will make it easier for you to plan your presentation. Identifying that intent is also one of the harder parts of planning a business presentation.
“A presentation is a voyage with purpose and it must be charted. The man who starts out going nowhere, generally gets there.” – Dale Carnegie
- Be absolutely clear on the intent of your presentation.
- Summarise your intent in one line
- Use your intent as your North Star to guide everything you say and how you say it.
Create a killer presentation. Step #4 – AIM part 3: What’s your one big take-away message?
Summary – your presentation needs a take-away message. this means one simple message so when someone asks “what was that presentation about” a listener can confidently answer what you want them to say., what typically goes wrong.
Many presentations have titles such as:
- “Quarterly strategy report”
- “Project X”
- “Manufacturing update”
- “Annual results”
These are all topics, not messages
Why does this matter? These titles do not help the audience. It only tells them something they already know. With a topic title you miss the opportunity of preparing your audience and getting them in the right mindset to be ready for your presentation.
So, instead, identify a message that summarises your presentation that you can use as its title. Keep improving the title until it properly captures what you want to say. For example:
- “Our strategy remains on track”
- “Launching Project X by December could double revenues next year”
- “Manufacturing: three problems we must address”
- “Profits up 5% this year despite Covid headwinds”
Then test your title on other people. Check if it generates the reaction you want.
“If you can’t write your idea on the back of my calling card, you don’t have a clear idea.” – David Belasco, theatre producer
- Decide the title of your presentation early.
- Check it generates the reaction you want.
- Use this to build the rest of your presentation.
- Re-test your message against A.I.M.
Create a killer presentation. Step #5 – What are the three parts of your presentation?
Summary – your brain is naturally tuned to hearing things in sets of three. if you can break your presentation into three parts that work together then it’s more likely to be successful..
What typically goes wrong? Many presentations are like shopping lists. The presentation covers multiple topics and jumps from one idea to the next.
What’s wrong with a shopping list approach? In the end, a huge amount of information has been transmitted but little has been received.
So, instead, remember that Less is More in a presentation. Help your audience by giving them a structure. A three part structure is one of the most useful planning shortcuts that you can use.
Once you are completely clear about your intent and your message, start developing a three part structure for your presentation. For example, if I wanted to give a presentation that shared advice on how to present, I would consider using one of the following structures:
Mistakes other people make / Tips you can use / How to become a great speaker or How to define your messages / How to structure your presentation / How to deliver your presentation or What bad looks like / what good looks like / what you can do differently
“ In writing and speaking, three is more satisfying than any other number. “ – Carmine Gallow, author
- Find your three part structure early.
- Use the structure to focus your efforts and guide your planning
Create a killer presentation. Step #6 – What is your story?
Summary – to improve your presentation skills you should underpin your presentation with a story or a narrative. no matter how dry your subject, by using a story it will be more memorable and more effective..
What typically goes wrong? As an expert, a typical presenter wants to share knowledge. For example, A few years ago, I helped a lawyer give a business presentation to investors. This audience was made up of private equity investors in businesses who also sat on their boards. The subject was the 2006 Companies Act and the Duties and Responsibilities of a Director. A dry subject.
In her first draft, she reviewed sections of the Act and highlighted problems that directors may face. For example, “Section 172 of the Act, sets out your overarching duties as a director. You must act in the way you consider, in good faith, would be most likely to promote the success of the company for the benefit of its members as a whole.”
As you can imagine, it was not the most exciting of business presentations.
Why does this matter? A precise business presentation may not be an interesting business presentation. And an uninteresting presentation will not be heard. Your job as a presenter is to make your business presentation interesting and easy for your audience.
What to do instead
What this means is that you need to find a story that fits what you want to talk about. This is key to improving presentation skills.
For example, for the investor director presentation above, we decided to title the presentation “How to keep your nose clean and yourself out of jail”
Then the presentation was based around a series of situations that anyone in the audience might face. She did not refer to any particular section of the Companies Act at all. Her text was:
“Imagine this situation. You turn up for a board meeting. You are a 10% shareholder and you are a director. At that board meeting the CEO announces that the company is near bankrupt and needs more funding. What should you do? Do you absent yourself, having a connected interest. Or do you declare your interest as a shareholder? Or do you carry as normal assuming business as usual?”
By framing it as a story you involve your audience and you make it easier for them to process what you say.
“Sometimes reality is too complex. Stories give it form.” – Jean Luc Godard, film director
- Find stories to tell, narratives to bring your facts to life.
- Tell the story behind the numbers.
- The dryer your subject matter, the more important stories become.
Create a killer presentation. Step #7 – Have you asked WHY?
Summary – ‘why’ comes before ‘what’ comes before ‘how’.
What typically goes wrong? When a speaker knows a subject well, it is easy for them to assume knowledge and talk about the nuances of what they know. I often describe this as the ‘How’ of a subject.
For example, when speaking about a new engine you have developed, you might say that we made the pistons more accurately, that you mix petrol more precisely and you have added a new technique of managing engine performance.
Why does this matter? This is one of the most common mistakes that experts make when giving presentations. They spend too much time explaining HOW something works, rather than explaining WHAT it is they are presenting about and WHY it is important.
Instead, to improve your presentation skills, phrase it like this, answering the WHY question: “We have designed a car engine that is more efficient and will get 100 miles to the gallon.”
“Start with Why” – Simon Sinek
- Check that you are clear why the audience will be interested.
- Imagine someone in the audience asking “What’s in it for me?” (WIIFM)
Learn about our executive presentation coaching courses
Create a killer presentation. Step #8 – Can you give a 90 second summary of your presentation?
Summary – the best way to test your planning and improve your presentation skills is by speaking a c 100-200 word summary of your presentation. this summary will test the rigour of your thinking, the clarity of your ideas and the robustness of your plan..
What typically goes wrong? The average poor presentation meanders from topic to topic and is more like a data dump than a well organised business presentation. It is rich in information but poor in story, structure and planning. It will not easily fit into a short sharp clear summary.
Why does this matter? Lack of planning = Lack of story = hard on your audience.
So, instead, Create a short summary of your presentation to test your thinking
- You can use it early in your planning to test your ideas.
- If you are working with colleagues you can share your thinking.
- If you are preparing a presentation for someone else, you can share your summary to test their reaction.
- If someone else is preparing your presentation, you can use a summary to check they are on track.
This is one of the most powerful of all shortcuts and will save you a huge amount of wasted time.
“If you can’t write your message in a sentence, you can’t say it in an hour.” – Dianna Booher, Author
- Test your ideas with a short summary.
- Use a critical audience.
- If it is not tight enough, keep refining your summary.
Summary – how to plan your presentation for success
- Use AIM as the first stage of preparing any presentation
- Answer the WHY questions in your presentation before the WHAT or HOW questions.
Transform your presentation skills with tailored coaching

We can help you present brilliantly. Thousands of people have benefitted from our tailored in-house coaching and advice – and we can help you too .
“I honestly thought it was the most valuable 3 hours I’ve spent with anyone in a long time.” Mick May, CEO, Blue Sky
For 15+ years we’ve been the trusted choice of leading businesses and executives throughout the UK, Europe and the Middle East to improve corporate presentations through presentation coaching, public speaking training and expert advice on pitching to investors.
Unlock your full potential and take your presentations to the next level with Benjamin Ball Associates.
Speak to Louise on +44 20 7018 0922 or email [email protected] to transform your speeches, pitches and presentations.
Or read another article..., how to improve your executive presence.
Learn how coaching can help you develop executive presence To succeed in business,…
How to Scale your Business – 8 Low Cost Ways
How do you scale a business? This is one of the big questions…
How to Make A Compelling Financial Presentation
Writing financial presentations is not easy. Typically, You have a lot of information…
How to Create a Winning Elevator Pitch – Top 11 tips
If you need to give a short pitch – an elevator pitch –…
Contact us for a chat about how we can help you with your presenting.
What leaders say about Benjamin Ball Associates
Ceo, plunkett uk.
"Thank you so much for an absolutely brilliant session yesterday! It was exactly what we were hoping for, and you did an incredibly job covering such a range of issues with 4 very different people in such short a session. It really was fantastic - thank you!"
James Alcock, Chief Executive, Plunkett UK
Manager, ubs.
"Essential if you are going to be a spokesperson for your business"
Senior Analyst, Sloane Robinson
"Being an effective communicator is essential to get your stock ideas across. This course is exactly what's needed to help you do just that!"
CEO, Blast! Films
“Our investment in the coaching has paid for itself many times over.”
Ed Coulthard
Corporate finance house.
“You address 95% of the issues in a quarter of the time of your competitor.”
Partner International
“Good insight and a great toolbox to improve on my presentations and delivery of messages to not only boards, analysts and shareholders but to all audiences”
CEO, Eurocamp
“We had a good story to tell, but you helped us deliver it more coherently and more positively.”
Steve Whitfield
Ceo, ipso ventures.
“Ben did a great job on our presentation. He transformed an ordinary set of slides into a great presentation with a clear message. Would definitely use him again and recommend him highly.”
Nick Rogers
“Moved our presentation into a different league and undoubtedly improved the outcome and offer we received.”
Let's talk about your presentation training needs
+44 20 7018 0922, [email protected], our bespoke presentation coaching services, investor pitch coaching, executive presentation coaching, public speaking training, executive media training, new business pitch coaching, privacy overview.
| Cookie | Duration | Description |
|---|---|---|
| cookielawinfo-checkbox-analytics | 11 months | This cookie is set by GDPR Cookie Consent plugin. The cookie is used to store the user consent for the cookies in the category "Analytics". |
| cookielawinfo-checkbox-functional | 11 months | The cookie is set by GDPR cookie consent to record the user consent for the cookies in the category "Functional". |
| cookielawinfo-checkbox-necessary | 11 months | This cookie is set by GDPR Cookie Consent plugin. The cookies is used to store the user consent for the cookies in the category "Necessary". |
| cookielawinfo-checkbox-others | 11 months | This cookie is set by GDPR Cookie Consent plugin. The cookie is used to store the user consent for the cookies in the category "Other. |
| cookielawinfo-checkbox-performance | 11 months | This cookie is set by GDPR Cookie Consent plugin. The cookie is used to store the user consent for the cookies in the category "Performance". |
| viewed_cookie_policy | 11 months | The cookie is set by the GDPR Cookie Consent plugin and is used to store whether or not user has consented to the use of cookies. It does not store any personal data. |
How to Give a Killer Presentation – Lessons from TED
On the Harvard Business Review website presenters can find a true gem: Lessons from Chris Anderson , curator of the TED conference series. While it was originally published in the magazine in 2013, I find this timeless insight for anybody who wants to improve presentation skills.
Chris starts out with a story to demonstrate that giving a good talk is highly coachable.
He recalls meeting Richard Turere, a 12-year old Masai boy, who came up with a solution to one of the biggest challenges livestock farmers in Kenya face: protecting their animals from lions and other wild animals. He devised a system of lights that created a sense of movement that scared off lions at night. The lights worked as imagined by Richard and soon villagers in other part of Kenya started installing Richard’s “lion lights”.
This is the kind of inspiring story that is perfect for a TED Talk . The only problem: Richard seemed not an ideal candidate to give a presentation to a large audience used to listening to polished speakers like Bill Gates and Al Gore. Richard was painfully shy and when he tried to explain his invention, the sentences came out incoherently. Despite of this, Chris invited Richard to tell his story at at the 2013 TED conference.
In the months prior to his presentation, the team at TED coached Richard to frame his story – to find the right place to begin, and to develop the story into a killer presentation.
Chris Anderson writes in the HBR article, “When he finally gave his talk at TED, in Long Beach, you could tell he was nervous, but that only made him more engaging — people were hanging on his every word. The confidence was there, and every time Richard smiled, the audience melted. When he finished, the response was instantaneous: a sustained standing ovation.”
So how does the TED team coach their presenters to frame, practice, and deliver their stories? The process to create a killer presentation starts six to nine months before the event. That’s right!
A real killer presentation requires lots of planning, devising, rehearsing, and lots of fine tuning along the way. The actual task of transforming a presentation from muddled to mesmerizing is a matter of hours…spread over a longer period of time.
How to Give a Killer Presentation Anywhere – Not Only at TED
Frame your story.
We all have good stories to tell. The most vital part of making your story compelling is to conceptualize it and put it in a frame that people want to hear.
Think about taking your audience on a journey. As with any journey, how you start it and where you finish it is vital to the entire experience. The most engaging speakers quickly introduce the topic, explain why they care so deeply about it, and convince the audience members that they should as well.
Don’t cram everything you know into your presentation. Instead, use a few specific examples to highlight your ideas. Your presentation won’t be rated by how broad it is, but rather by how deep you can involve your listener into a few well explained details about the problem you are solving.
Plan Your Delivery
There are three main ways to give a talk. You can read it off a script or teleprompter. You can develop a set of bullet points that map out what you’re going to cover in each section. Or you can memorize your talk to deliver it word for word as you have practiced it.
Of course, the last method is the most time consuming during your preparation. It will require hours of rehearsing. Memorizing your talk, however, is also the most effective once you are in front of your audience. It is the one the TED team recommends to their presenters for a good reason: many of their best and most popular talks have been memorized word for word.
What if you don’t have the time or the presentation is not as significant as a TED Talk with 1,400 people in the audience and millions behind their computer screens? Go with bullet points on flash cards. As long as you know what you want to say to each point and transition well from section to section, you’ll be fine.
Another point Chris makes in the HBR article is to sound natural. Present your information and story in a conversational tone. Don’t orate, don’t lecture. Be as natural as if you would talk to a group of your friends.
Develop Stage Presence
Just the thought of standing in front of an audience can be extremely frightening. To take off some of this edge, always remember that your words, story, and substance is much more important to the audience than the way you stand or if you are visibly nervous.
Richard seems quite nervous in his presentation about the “lion lights”, but people in the audience didn’t care at all about that. In fact, he built better rapport because the audience members could identify with this nervousness. Furthermore, they sensed that he is confident talking about his experience and story. That’s one of the reasons his talk was receiving such a positive feedback.
One of the biggest mistakes the TED team sees in early rehearsals is that people move their bodies too much. They sway from side to side or shift their weight from one leg to the other. They coach their presenters to keep their lower bodies motionless, which can dramatically improve stage presence. Although there are some presenters that are able to walk around the stage during their presentation (the late Steve Jobs was very natural doing it), the majority of presenters is better off standing still and relying on hand gestures and facial expressions.
The one aspect that will do most for your successful presentation is eye contact. Pick a few audience members and imagine them as friends you haven’t seen in a while. Make solid eye contact with them while you update them on your work.
Plan the Multimedia
You probably have heard the advice about PowerPoint (or similar presentation software): Keep it Simple! Don’t use your presentation deck as a crutch, as a substitute for notes. And never, ever read from your slides. It will only make the audience think that they could do that themselves in the comfort of their home or office. Instead, use powerful images that visualize the key points of your presentation.
In Richard’s presentation about his “lion lights” there are no text slides. They are not needed. There are, however, many photographs that engage the audience to take a short journey into Richard’s world.
Putting it Together
The TED team starts helping their presenters at least six months in advance so that they’ll have plenty of time to practice. They want people’s talks in final form one month prior to the event. Why? The more practice the presenters can put in during the final weeks, the more compelling the presentation will be.
The most important thing to keep in mind though is that presentations rise and fall on the quality of ideas, the narrative, and the passion of the speaker. It’s about substance, not speaking style or multimedia pyrotechnics. If you have something to say, you can build a great talk.
Remember, there is no one good way to give a killer presentation. The most memorable talks offer something fresh, engaging, and relevant.
Read the full article on the Harvard Business Review website.
Get instant access to my newsletter where I share my best presentation tips.
100% Privacy. No Spam.
More Presentation Tips

Virtual Card Magic for Online Presentations
Virtual Card Magic is a ridiculously cool card trick designed specifically for online presentations and meetings.

10 Key Takeaways from a Shark Tank Success Story
Learn from Haley McClain Hill’s Shark Tank success story what it takes to prepare for Shark Tank — and secure a deal!

Enter the Spotlight with PowerPoint Cameo
PowerPoint Cameo is a hidden gem, a powerful feature that allows you to seamlessly integrate your live video feed into your presentation slides.
Latest PitchZone Episode
Related posts.

ChatGPT Voice – A Translator in Your Pocket
For anyone who has ever wished for a universal translator à la ‘Star Trek’ – our sci-fi dreams are coming closer to reality, thanks to ChatGPT Voice.

Create Beautiful PowerPoint Slides with ChatGPT and VBA
Here’s a quick tip to boost your productivity by letting ChatGPT and VBA do the heavy lifting when creating PowerPoint slide decks.
Copyright 2024 © All rights Reserved.
Privacy policy.
How to Give a Killer Presentation: 18 Top Tips

People attend presentations to learn and gain useful insight. But way too often, we see the audience yawn, scroll on their phones or check their watch, wishing that the time would go faster.
Is it that the content of the presentation isn’t interesting enough, or that the speaker doesn’t know how to engage the audience?
Whatever the reason, delivering an engaging presentation is an art that takes some time to master.
Based on my own experience from the stage, and from observing other speakers at industry-leading conferences, I’ve collected these 18 top tips. May they help you give a presentation that will wow your audience:
- Plan your storyline
- Use the rule of three
- Simplify your slides
- Include numbers
- Use the power of visuals
- Practice relentlessly
- Greet the audience in their local language
- Break the ice at the start
- Engage your audience with live polls
- Move around the stage and make gestures
- Smile and make eye contact
- Consider using props
- Go among the audience
- Give rewards for participation
- Prompt a discussion in the audience
- Build in time for Q&A
- Crowdsource questions from the audience
- Gather feedback
1. Plan your storyline
A powerful story can make your whole presentation. Take TED talks, for instance. They’re all based on captivating stories that support the main argument or line of thought of each speech.
Give your presentation a concept. Use a classic narrative structure, from a gripping outset to an impressive end. A presentation designer Nancy Duarte advises presenters to spend twice as much time on framing the storyline than creating the actual slides.
Also, don’t forget to add emotional details and power words. These will make your audience feel much more connected to you. People will eventually forget your slides and your presentation, but they will not forget how you made them feel.
2. Use the rule of three
People can usually remember only three main points from presentations, so take advantage of this psychological phenomenon.
While creating your storyline, think of three key messages that you want your audience to walk out of the room with. To make these three key points stick, you need to make them short, memorable and attention-grabbing.
On the other hand, if your presentation revolves around one main argument, make use of the Aristotelian “triptych” method: “Tell them what you’re going to tell them. Tell them. Then tell them what you told them.” In a nutshell, you should properly introduce the point you will be making, then make your point, and then wrap up with summarizing the main point.
3. Simplify your slides
No matter how rich in content your slides are, if they’re too crowded, nobody is going to read them. Too much text on slides actually takes your audience’s attention away from your presentation, which hinders the learning process.
Make your slides as simple as possible and try to present only one idea per slide. Sometimes, one powerful sentence, a number, or even one word, can say more than a slide full of bullet points.
4. Include numbers
When used sensibly, numbers can strengthen your point and back up your arguments. To make data easy for your audience to digest, you need to make it specific, relevant and contextual.
When Steve Jobs introduced the first iPod , he did not emphasize its 5GB storage and 185g weight. Instead, he repeatedly said that it could hold 1,000 songs and physically manifested that he could fit it into his pocket. This number was easy for the audience to remember, and called even more attention to its tiny size.
5. Use the power of visuals
Videos or images not only engage the audience but also help to evoke emotions that are otherwise super difficult for speakers to elicit by themselves.
Make sure that the visuals you use support the main point of your presentation, or demonstrate what you’re talking about. This works very well in the creative industry, where visual aids are often necessary to complement the main content.
However, if you’re going to use video, be cautious. A too lengthy or unengaging video may put people to sleep rather than keep them attentive.
6. Practice relentlessly
Attending a presentation where the speaker keeps looking down at his notes is painful so don’t underestimate this point. For instance, Winston Churchill rehearsed for hours, even days, to deliver a 10-minute long speech.
Memorize your presentation flow by heart if need be. Do it to the extent that you won’t need the notes.
During your rehearsals, use a video camera to record yourself in order to see where you stutter, where you seem nervous and how you work with your body language. Don’t be afraid to ask a friend to give you feedback.
Tim Ferris , the author of The 4-Hour Workweek, follows a Spartan’s preparation for his public speeches. He splits his presentation into several segments and he goes through each one of them up to ten times.
7. Greet your audience in their local language
As a speaker, you often find yourself addressing an international audience, whether it is at a big conference or an internal company meeting joined by remote teams.
Greeting international participants in their local language gives a nice personal touch to the offset of your presentation. It helps you create a connection and the feeling of intimacy with the people sitting before you.
I always memorize how to say “Hello” and “How are you?” in the local language, and use them as soon as I come onstage. You can even take it a step further and adjust your presentation ad hoc to the audience, by making local references.
For example, Google’s Digital Marketing Evangelist, Avinash Kaushik , started his talk at the Marketing festival by showing pictures from his tour around the hosting city of Brno, Czech Republic. Moreover, he used the Czech websites that the audience was closely familiar with, instead of international ones, to get his point across.
8. Break the ice at the start
Hook your audience right off the bat. Using an effective icebreaker will help you set the stage and energize your attendees.
Here’s an inspiration for you: At the 2018 World Education Congress (WEC) , I asked people to close their eyes and think of a presentation session that had recently impressed them. After 30 seconds, I invited them to share their dream session with their neighbor and describe it using one word, before submitting it to a Slido word cloud poll.

Next, I asked them to picture the usual experience of attending a presentation and describe it again, using a single word. Seeing the differences in the two consequent word cloud polls was very thought-provoking and sparked up a discussion among the attendees.

Other than using technology, you can liven up your audience with a classic show of hands or other brisk icebreaking activities , such as rock, paper, scissors or live barometer.
Related story: The Complete List of 200 Icebreaker Questions and Tips On How to Use Them
9. Engage your audience with live polls
Once you win your audience over, keep up the pace by creating enough interaction points throughout your presentation.
Live polling is your best bet here. When smartly used, live polls will keep your attendees engaged during the whole length of your presentation. They also help you to effectively collect your audience’s insights, which you can then showcase on screen. This multiplies the learning element of your presentation.
In general, I follow the human attention span and use a poll every 8-10 minutes, which is 5-6 polls for a 60-minute talk, maximum.
To give you an example, during the latest webinar I led, I asked the participants a simple rating poll: “How would you rate interaction in the classroom today?”
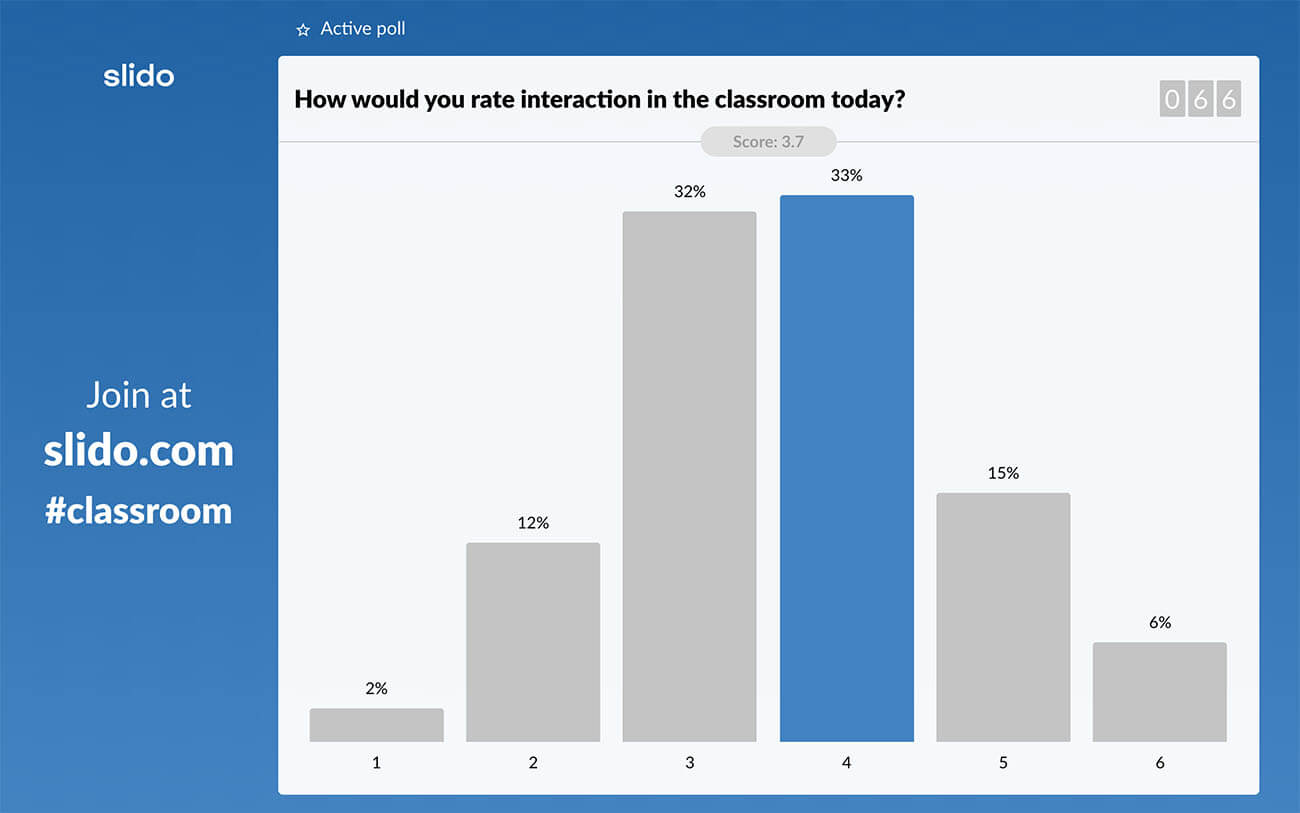
The results set a good ground for the main argument I was going to make about insufficient interactivity in education, and really helped me make my point.
On top of that, this strategy allows me to break the long content deliveries into more digestible chunks, regain the audience’s attention, and ignite conversations based on the results.
The last point is particularly important. Live polls make sense only when you facilitate their use. So make sure to always follow up on the results, share your thoughts on them, or get the audience to share why they voted the way they did.
Related story: The Complete Guide: How to Use and Facilitate Slido Polls in Your Presentation
10. Move around the stage and make gestures
If you stand rigidly in one spot or behind a speaker’s stand, you will only appear unconfident and nervous. Think of yourself as an actor on stage, and your presentation as your performance. Use open, big gestures, point in the direction of the audience, or slowly walk about the stage.
At this year’s Festival of Marketing , Mark Ritson – who was opening the event – reminded me of the importance of using body language. He kept pacing the stage in a natural way and was gesturing throughout the entire length of his speech. It was definitely one of the most engaging sessions I’ve attended this year.
11. Smile and make eye contact
The way you communicate with your audience through your facial expressions makes a huge difference. So don’t look down at your notes, don’t look at your slides, but keep your eyes set on your audience.
Lisa Wentz, a public speaking expert, advises to pick 3 people in the audience that you like, each one at a different corner of the auditorium, and make eye contact with them throughout your presentation. However, avoid staring at one person for too long. Use the selected people only as navigation points that will help you scan the room.
12. Consider using props
Demonstrating the point with the use of props is a powerful way to help the attendees visualize what is being described verbally. Showing a prop at the right moment can help you catch your audience’s attention and enforce your story.
Neuroscientist Jill Bolte Taylor brought a real human brain on stage during her emotional TED talk to explain what had happened to her when she had a stroke. She touched the audience with this demonstration and left them in complete awe.
13. Go among the audience
Asking people questions may feel impersonal if you stand onstage. On top of that, large auditoriums often make it difficult to create intimacy with your audience.
Draw inspiration from rock singers here and “jump” off the stage. Going among the audience will help you build a stronger bond with them and your presentation will feel more personal.
This approach is invaluable if you hope to collect impromptu answers after you have asked your question. Move slowly around the room, and when someone shuffles or raises a hand, approach them with a mic and elicit an answer.
When another hand shoots up, move to that corner of the room, and so on. The point here is to be as close to your audience as possible.
If possible, check the room advance to get used to the space arrangements. This will help you move around more naturally and with more confidence.
14. Give rewards for participation
Despite all your efforts, the audience might need a bit of a nudge. Giving out small rewards can bring another interactive element to your presentation. You can go with the event merchandise or small treats, like chocolates and candy.
For example, at the Eventex conference , one of the speakers, Victor Neyndorff , encouraged people to share their thoughts by handing out chocolate from the Netherlands, his home country.
To give you another idea, at the Jam London conference , the organizers decided to give away books to those attendees who were the most active in asking questions via Slido. This really helped incentivize the audience to participate and improved the dialogue in the room.
15. Prompt a discussion in the audience
You can give audience engagement another spin by giving your attendees an activity that they can participate in.
For example, you can present a statement for the participants to discuss, or give them a task to solve in groups. Where appropriate, walk around the room, join the conversations, and encourage people to talk to each other.
At the Conventa Crossover conference in Slovenia, moderator Jan-Jaap In der Maur put people in small groups and asked them to share the technological trends that they believed will have the biggest impact on the industry in the near future.
Then he collected a few comments from the floor to open a discussion with the whole room.
Simple. Engaging. Useful.
If facilitated properly, activities like these can work equally well with an audience of 20 people as they can with 2,000.
Related story: 5 Essential Pieces of The Audience Engagement Puzzle
16. Build in time for the Q&A
Even if you incorporate interactive elements to your presentation, your audience will surely have additional questions.
For that reason, don’t be scared to allocate as much as 10-20 minutes to the Q&A, depending on the length of your presentation slot.

After I finish my talk, instead of asking, “Are there any questions?” (which typically leads to silence), I like to ask, “What are your questions?”, or say, “Now, let’s get to your questions.” In case I don’t get an instant reaction from the audience, I get off the stage and walk among the audience to encourage the discussion.
In rare moments when no questions come up, I kick off the Q&A by saying: “What people usually ask me is…” and then give an answer. In 9/10 times, the discussion catches on.
17. Crowdsource questions from the audience
Lack of audience questions doesn’t necessarily mean that your audience doesn’t have questions. They may just be uncomfortable with speaking up in public.
Live Q&A tools like Slido allow you to effortlessly crowdsource questions from your audience throughout your presentation via an app.
Compared to passing the mic amongst the people in the audience, you will give everyone an equal chance to ask questions, regardless of their level of shyness.
If you’re using a Q&A app, it’s important that you introduce and facilitate it properly. I often say something like: “Take a minute and think about what you’ve just heard. Come up with a question that you have, and submit it to Slido.” It works every single time.
Then, just take a look at the screen, or a confidence monitor, and address the questions that have the most upvotes.
If you display the crowdsourced questions on the screen, read each question out loud when addressing it. It will help your audience – even the ones sitting at the back – to know which question you are answering.
Extra tip: Sometimes, you get way more questions from your audience than you can answer during your time-limited Q&A slot. Don’t leave them hanging in the air. Here you’ll find 5 tips on what to do with unanswered questions after your Q&A .
18. Gather feedback
Feedback is priceless for improving your presentation skills. There’s never enough of it. You can collect feedback easily via Slido feedback survey . Combine rating polls for quick assessment and open text polls to give your participants space for more in-depth comments.
Your feedback survey could look something like this:
- How would you rate this presentation? (rating poll)
- What is your main takeaway from this session? (open text poll)
- What would you improve? (open text poll)
To boost the response rate, make sure that you ask your attendees to fill out the survey while they’re still in the room.
With the tips I’ve listed above, you’ll be able to turn your presentation or lecture from a one-way content broadcast into an exciting conversation between you and your attendees.
Engage your attendees with Slido live Q&A and polls.
Try Slido now
Get just a single email per month with our best articles.

Presentations
5 ways to use slido in google slides.
In this article, you’ll find examples of poll questions and quizzes that you can create with Slido and use in...

7 Interactive Poll Ideas for Your Next PowerPoint Presentation
Looking for new ways to make your PowerPoint presentation more interactive? Try live polls. With polls, you can collect non-verbal...

How to Give an Interactive PowerPoint Presentation in 2023
Presenting online is tough, yes. You can’t really connect with your audience. You often don’t even know whether those mute...
We use essential cookies to make Venngage work. By clicking “Accept All Cookies”, you agree to the storing of cookies on your device to enhance site navigation, analyze site usage, and assist in our marketing efforts.
Manage Cookies
Cookies and similar technologies collect certain information about how you’re using our website. Some of them are essential, and without them you wouldn’t be able to use Venngage. But others are optional, and you get to choose whether we use them or not.
Strictly Necessary Cookies
These cookies are always on, as they’re essential for making Venngage work, and making it safe. Without these cookies, services you’ve asked for can’t be provided.
Show cookie providers
- Google Login
Functionality Cookies
These cookies help us provide enhanced functionality and personalisation, and remember your settings. They may be set by us or by third party providers.
Performance Cookies
These cookies help us analyze how many people are using Venngage, where they come from and how they're using it. If you opt out of these cookies, we can’t get feedback to make Venngage better for you and all our users.
- Google Analytics
Targeting Cookies
These cookies are set by our advertising partners to track your activity and show you relevant Venngage ads on other sites as you browse the internet.
- Google Tag Manager
- Infographics
- Daily Infographics
- Popular Templates
- Accessibility
- Graphic Design
- Graphs and Charts
- Data Visualization
- Human Resources
- Beginner Guides
Blog Beginner Guides How To Make a Good Presentation [A Complete Guide]
How To Make a Good Presentation [A Complete Guide]
Written by: Krystle Wong Jul 20, 2023

A top-notch presentation possesses the power to drive action. From winning stakeholders over and conveying a powerful message to securing funding — your secret weapon lies within the realm of creating an effective presentation .
Being an excellent presenter isn’t confined to the boardroom. Whether you’re delivering a presentation at work, pursuing an academic career, involved in a non-profit organization or even a student, nailing the presentation game is a game-changer.
In this article, I’ll cover the top qualities of compelling presentations and walk you through a step-by-step guide on how to give a good presentation. Here’s a little tip to kick things off: for a headstart, check out Venngage’s collection of free presentation templates . They are fully customizable, and the best part is you don’t need professional design skills to make them shine!
These valuable presentation tips cater to individuals from diverse professional backgrounds, encompassing business professionals, sales and marketing teams, educators, trainers, students, researchers, non-profit organizations, public speakers and presenters.
No matter your field or role, these tips for presenting will equip you with the skills to deliver effective presentations that leave a lasting impression on any audience.
Click to jump ahead:
What are the 10 qualities of a good presentation?
Step-by-step guide on how to prepare an effective presentation, 9 effective techniques to deliver a memorable presentation, faqs on making a good presentation, how to create a presentation with venngage in 5 steps.
When it comes to giving an engaging presentation that leaves a lasting impression, it’s not just about the content — it’s also about how you deliver it. Wondering what makes a good presentation? Well, the best presentations I’ve seen consistently exhibit these 10 qualities:
1. Clear structure
No one likes to get lost in a maze of information. Organize your thoughts into a logical flow, complete with an introduction, main points and a solid conclusion. A structured presentation helps your audience follow along effortlessly, leaving them with a sense of satisfaction at the end.
Regardless of your presentation style , a quality presentation starts with a clear roadmap. Browse through Venngage’s template library and select a presentation template that aligns with your content and presentation goals. Here’s a good presentation example template with a logical layout that includes sections for the introduction, main points, supporting information and a conclusion:

2. Engaging opening
Hook your audience right from the start with an attention-grabbing statement, a fascinating question or maybe even a captivating anecdote. Set the stage for a killer presentation!
The opening moments of your presentation hold immense power – check out these 15 ways to start a presentation to set the stage and captivate your audience.
3. Relevant content
Make sure your content aligns with their interests and needs. Your audience is there for a reason, and that’s to get valuable insights. Avoid fluff and get straight to the point, your audience will be genuinely excited.
4. Effective visual aids
Picture this: a slide with walls of text and tiny charts, yawn! Visual aids should be just that—aiding your presentation. Opt for clear and visually appealing slides, engaging images and informative charts that add value and help reinforce your message.
With Venngage, visualizing data takes no effort at all. You can import data from CSV or Google Sheets seamlessly and create stunning charts, graphs and icon stories effortlessly to showcase your data in a captivating and impactful way.

5. Clear and concise communication
Keep your language simple, and avoid jargon or complicated terms. Communicate your ideas clearly, so your audience can easily grasp and retain the information being conveyed. This can prevent confusion and enhance the overall effectiveness of the message.
6. Engaging delivery
Spice up your presentation with a sprinkle of enthusiasm! Maintain eye contact, use expressive gestures and vary your tone of voice to keep your audience glued to the edge of their seats. A touch of charisma goes a long way!
7. Interaction and audience engagement
Turn your presentation into an interactive experience — encourage questions, foster discussions and maybe even throw in a fun activity. Engaged audiences are more likely to remember and embrace your message.
Transform your slides into an interactive presentation with Venngage’s dynamic features like pop-ups, clickable icons and animated elements. Engage your audience with interactive content that lets them explore and interact with your presentation for a truly immersive experience.

8. Effective storytelling
Who doesn’t love a good story? Weaving relevant anecdotes, case studies or even a personal story into your presentation can captivate your audience and create a lasting impact. Stories build connections and make your message memorable.
A great presentation background is also essential as it sets the tone, creates visual interest and reinforces your message. Enhance the overall aesthetics of your presentation with these 15 presentation background examples and captivate your audience’s attention.
9. Well-timed pacing
Pace your presentation thoughtfully with well-designed presentation slides, neither rushing through nor dragging it out. Respect your audience’s time and ensure you cover all the essential points without losing their interest.
10. Strong conclusion
Last impressions linger! Summarize your main points and leave your audience with a clear takeaway. End your presentation with a bang , a call to action or an inspiring thought that resonates long after the conclusion.
In-person presentations aside, acing a virtual presentation is of paramount importance in today’s digital world. Check out this guide to learn how you can adapt your in-person presentations into virtual presentations .

Preparing an effective presentation starts with laying a strong foundation that goes beyond just creating slides and notes. One of the quickest and best ways to make a presentation would be with the help of a good presentation software .
Otherwise, let me walk you to how to prepare for a presentation step by step and unlock the secrets of crafting a professional presentation that sets you apart.
1. Understand the audience and their needs
Before you dive into preparing your masterpiece, take a moment to get to know your target audience. Tailor your presentation to meet their needs and expectations , and you’ll have them hooked from the start!
2. Conduct thorough research on the topic
Time to hit the books (or the internet)! Don’t skimp on the research with your presentation materials — dive deep into the subject matter and gather valuable insights . The more you know, the more confident you’ll feel in delivering your presentation.
3. Organize the content with a clear structure
No one wants to stumble through a chaotic mess of information. Outline your presentation with a clear and logical flow. Start with a captivating introduction, follow up with main points that build on each other and wrap it up with a powerful conclusion that leaves a lasting impression.
Delivering an effective business presentation hinges on captivating your audience, and Venngage’s professionally designed business presentation templates are tailor-made for this purpose. With thoughtfully structured layouts, these templates enhance your message’s clarity and coherence, ensuring a memorable and engaging experience for your audience members.
Don’t want to build your presentation layout from scratch? pick from these 5 foolproof presentation layout ideas that won’t go wrong.

4. Develop visually appealing and supportive visual aids
Spice up your presentation with eye-catching visuals! Create slides that complement your message, not overshadow it. Remember, a picture is worth a thousand words, but that doesn’t mean you need to overload your slides with text.
Well-chosen designs create a cohesive and professional look, capturing your audience’s attention and enhancing the overall effectiveness of your message. Here’s a list of carefully curated PowerPoint presentation templates and great background graphics that will significantly influence the visual appeal and engagement of your presentation.
5. Practice, practice and practice
Practice makes perfect — rehearse your presentation and arrive early to your presentation to help overcome stage fright. Familiarity with your material will boost your presentation skills and help you handle curveballs with ease.
6. Seek feedback and make necessary adjustments
Don’t be afraid to ask for help and seek feedback from friends and colleagues. Constructive criticism can help you identify blind spots and fine-tune your presentation to perfection.
With Venngage’s real-time collaboration feature , receiving feedback and editing your presentation is a seamless process. Group members can access and work on the presentation simultaneously and edit content side by side in real-time. Changes will be reflected immediately to the entire team, promoting seamless teamwork.

7. Prepare for potential technical or logistical issues
Prepare for the unexpected by checking your equipment, internet connection and any other potential hiccups. If you’re worried that you’ll miss out on any important points, you could always have note cards prepared. Remember to remain focused and rehearse potential answers to anticipated questions.
8. Fine-tune and polish your presentation
As the big day approaches, give your presentation one last shine. Review your talking points, practice how to present a presentation and make any final tweaks. Deep breaths — you’re on the brink of delivering a successful presentation!
In competitive environments, persuasive presentations set individuals and organizations apart. To brush up on your presentation skills, read these guides on how to make a persuasive presentation and tips to presenting effectively .

Whether you’re an experienced presenter or a novice, the right techniques will let your presentation skills soar to new heights!
From public speaking hacks to interactive elements and storytelling prowess, these 9 effective presentation techniques will empower you to leave a lasting impression on your audience and make your presentations unforgettable.
1. Confidence and positive body language
Positive body language instantly captivates your audience, making them believe in your message as much as you do. Strengthen your stage presence and own that stage like it’s your second home! Stand tall, shoulders back and exude confidence.
2. Eye contact with the audience
Break down that invisible barrier and connect with your audience through their eyes. Maintaining eye contact when giving a presentation builds trust and shows that you’re present and engaged with them.
3. Effective use of hand gestures and movement
A little movement goes a long way! Emphasize key points with purposeful gestures and don’t be afraid to walk around the stage. Your energy will be contagious!
4. Utilize storytelling techniques
Weave the magic of storytelling into your presentation. Share relatable anecdotes, inspiring success stories or even personal experiences that tug at the heartstrings of your audience. Adjust your pitch, pace and volume to match the emotions and intensity of the story. Varying your speaking voice adds depth and enhances your stage presence.

5. Incorporate multimedia elements
Spice up your presentation with a dash of visual pizzazz! Use slides, images and video clips to add depth and clarity to your message. Just remember, less is more—don’t overwhelm them with information overload.
Turn your presentations into an interactive party! Involve your audience with questions, polls or group activities. When they actively participate, they become invested in your presentation’s success. Bring your design to life with animated elements. Venngage allows you to apply animations to icons, images and text to create dynamic and engaging visual content.
6. Utilize humor strategically
Laughter is the best medicine—and a fantastic presentation enhancer! A well-placed joke or lighthearted moment can break the ice and create a warm atmosphere , making your audience more receptive to your message.
7. Practice active listening and respond to feedback
Be attentive to your audience’s reactions and feedback. If they have questions or concerns, address them with genuine interest and respect. Your responsiveness builds rapport and shows that you genuinely care about their experience.

8. Apply the 10-20-30 rule
Apply the 10-20-30 presentation rule and keep it short, sweet and impactful! Stick to ten slides, deliver your presentation within 20 minutes and use a 30-point font to ensure clarity and focus. Less is more, and your audience will thank you for it!
9. Implement the 5-5-5 rule
Simplicity is key. Limit each slide to five bullet points, with only five words per bullet point and allow each slide to remain visible for about five seconds. This rule keeps your presentation concise and prevents information overload.
Simple presentations are more engaging because they are easier to follow. Summarize your presentations and keep them simple with Venngage’s gallery of simple presentation templates and ensure that your message is delivered effectively across your audience.

1. How to start a presentation?
To kick off your presentation effectively, begin with an attention-grabbing statement or a powerful quote. Introduce yourself, establish credibility and clearly state the purpose and relevance of your presentation.
2. How to end a presentation?
For a strong conclusion, summarize your talking points and key takeaways. End with a compelling call to action or a thought-provoking question and remember to thank your audience and invite any final questions or interactions.
3. How to make a presentation interactive?
To make your presentation interactive, encourage questions and discussion throughout your talk. Utilize multimedia elements like videos or images and consider including polls, quizzes or group activities to actively involve your audience.
In need of inspiration for your next presentation? I’ve got your back! Pick from these 120+ presentation ideas, topics and examples to get started.
Creating a stunning presentation with Venngage is a breeze with our user-friendly drag-and-drop editor and professionally designed templates for all your communication needs.
Here’s how to make a presentation in just 5 simple steps with the help of Venngage:
Step 1: Sign up for Venngage for free using your email, Gmail or Facebook account or simply log in to access your account.
Step 2: Pick a design from our selection of free presentation templates (they’re all created by our expert in-house designers).
Step 3: Make the template your own by customizing it to fit your content and branding. With Venngage’s intuitive drag-and-drop editor, you can easily modify text, change colors and adjust the layout to create a unique and eye-catching design.
Step 4: Elevate your presentation by incorporating captivating visuals. You can upload your images or choose from Venngage’s vast library of high-quality photos, icons and illustrations.
Step 5: Upgrade to a premium or business account to export your presentation in PDF and print it for in-person presentations or share it digitally for free!
By following these five simple steps, you’ll have a professionally designed and visually engaging presentation ready in no time. With Venngage’s user-friendly platform, your presentation is sure to make a lasting impression. So, let your creativity flow and get ready to shine in your next presentation!
Discover popular designs

Infographic maker

Brochure maker

White paper online

Newsletter creator

Flyer maker
Timeline maker

Letterhead maker
Mind map maker

Ebook maker
10 Strong Opening Slides to Start A Presentation (With Examples!)
Hrideep barot.
- Presentation

It is weird how now that we all live on our laptops and depend on them for entertainment and livelihood, things that shock us aren’t that many. This is a big itchy spot when it comes to engaging audiences and having a killer opening.
Lets focus on getting that perfect first slide in your presentation to help you kick off on the right foot.
How to open presentations
Opening presentations is an extremely daunting task. The worry of putting your best foot forward but at the same time not coming across as arrogant and the whole issue with fanning your armpits just before you step on the stage.
Yeah, I know. I relate, I think most of us do. We have braved those sweat patches and we have conquered.
It is time to up our opening game and while I will be getting to the ways we can do that, you can also check out this video for a quick idea.
What should be the first slide of a presentation?
Your first slide, needs to be impactful, with minimal content. An extremely difficult balance to maintain, but! Not impossible.
Your first slide, traditionally, is your name, the topic you are going to speak on and maybe on or two other details with MAYBE an image or some other graphics.
Gone are the days when we open speeches or presentations the traditional way, nothing wrong with it, but doing something “not normal” often helps us get people’s attention and that is the easiest way to get your points across and have them received positively.
Let’s check out a few ways you can open slides for a strong opening!
Strong Opening Slide Ideas
We’ve got our thinking cap on, let’s get cracking!
There are so many ways we can have a strong opening, even when you think presentations limit you.
Think of it this way, because people know you’re going to presenting something, they are going to give you full control of a projector. A big ass screen for all to see. If that isn’t filled with potential, I don’t know what is.
Well, with great power comes great responsibility, so let’s check out a few ways we can have killer opening slides , while of course being responsible… ish.
Idea 1: Introduction
There is no better way to get the audience to remember you than putting a giant photo of yourself on the screen and going, this is me, – an extremely edited version of me, but still, me. 🙂
Buddy. No. That was an attempt at being the funny – clever person. Clearly it didn’t work.
Don’t get me wrong, talking about yourself is good, important even to some extent, but that is it you see, it isn’t the fact that you’re talking about yourself that is the problem but what are you talking about that is.
The usual go to is to list out your biodata for the audience to read on the screen, while you speak the exact same thing off of the presentation. This is where we go wrong, no one wants to know about all your seven Ph.Ds. Bruce. (get the reference please)
Be proud of your qualifications, you earned them, but know when and where which qualification might be useful.
For example, you are a certified chartered accountant and have written plays that were on Broadway. In a screenwriting workshop / panel / seminar, as great as getting your chartered accountancy is, your experience as a writer holds way more value and is what will help you get the audience’s attention.
Let’s create an opening slide with the above example.

I used these polaroid photo ideas because for a play on Broadway, we’d love to see pictures! You can use tasteful pictures and even stock photos to help your audience get the right idea of your background.
Of course, I used these random paper elements to give it a more “writer” feel and also because this is my aesthetic, but you need to remember that this is your presentation and no cookie cutter mould will work. Even templates are meant to be edited to suit your needs.
Idea 2: Quiz
Is this to make your audience feel dumb? As much as that chaotic evil side of you may want to. Never do that. Respect their experiences as much as you would want them to respect yours.
Starting off with a quiz is a great way to warm up the crowd and get them involved in your presentation. Give them something to think about and it honestly doesn’t matter if they get it right or wrong, what matters is that they are trying to answer and interact!

Quizzes are a great ice breaker and also a great tool to get the audience going, you can also try to have a one off question or a series of questions.
Lets take the slide as an example, it could be for a presentation on a film industry and the question could be, guess the film from these three pictures, or they could be three different questions.
Remember as an opening slide, it should neither be text nor image heavy, just the right amount.
You could even create a game out of those quizzes and have checked off your list and even use these as a starting off point and come back to the topics (which could be your answers) while using this quiz as a reference point. The possibilities are endless!
Idea 3: Stimulation of Imagination
It always great to know what your audience is thinking, or in the least get them thinking!
You see, once they start thinking, they begin forming an opinion about the topic, which gets them invested and since you are the person addressing the topic, they will begin comparing their point of view / opinion with what they are saying.
There will always be different perspectives, what matters here is that they are invested enough to pay attention to you.
A really easy way to help them get started with forming an opinion is, asking them to take a minute to think about something.
For example: Think about a dancing monkey.
Can some of you describe the monkey you imagined, in the comment section? Was it wearing tap shoes and a top hat? Was it wearing a marching band uniform? Did it have your best friend’s face on it? Mine did!
Each of you had your own Dancing Monkey, and if thinking about it for a few seconds made it your own, imagine the attachment you can build by just spending a few minutes or even the duration of a presentation on it!

For example, you’re taking a presentation on perspectives or psychology. You can display this image and ask them what they think of it. Some may think about freedom, some loneliness and some people’s thoughts may be so profound that we could’ve never thought of it!
Idea 4: Video
This could work just as marvellous as sharing an image and opening a short discussion on its interpretations. You could even start with a video and use it as a segue into your presentation.
For example this video could be used as a great example for a marketing strategy by the brand and could be a great way to get the audience interested given the emotional quotient and relatable sibling content.
Idea 5: Image
Using an image might not necessarily mean that you can only invite the audience to imagine and think on their own. You can use an image to start your presentation and help get your point across.

You see that how the image is the hero of the slide? There is text, definitely, but much smaller, it looks as a complementary to the image instead of the other way around.
In this slide for example, assume poverty is the topic, a very telling image of poverty could help get the conversation started and make the audience more receptive of the topic.
An image in a way helps them “put a face” to the issue and that makes is easier for you to hold their attention and keep it.
Idea 6: Quote
It is well known and understood how impactful the right quote at the right time can be.
Lets focus on some things that people can often get wrong when using quotes.
Firstly, using long quotes, this is a no no when it comes to presentations because, then the audience will be in a rush to read the whole quote and if your point is made before then, well, we won’t get the desired effect will we?
Another thing to keep in mind is to not have a quote just to use it as a quote, pretty cryptic, honestly it is simple, if you are giving a presentation on a person and using their quote or you are using a random quote, make sure to have something to add to it.
It could be something simple. For example when talking about a person’s life:
“When this person said this, they were on their death bed, but they had lead a vivacious life until then to say the least, let’s start at the very beginning…”

Notice how despite there being a background picture, a text box, a bird in the corner, and all that, the text is what is the hero of the slide. You could even add a picture of the person whom you are quoting if it seems relevant.
Remember to always give credit where it is due. It never hurts.
Idea 7: Story
Who doesn’t love a good story? Storytelling is a major part of public speaking where animation, emotion and gestures and tones play a huge role in delivering your point.
With presentations, you need to remember to not just select any story, you need find / write a story that connects well to your topic, for example, if we are speaking about technology, a story about Alice and her looking glass don’t really give you much room to work in a segue.
Storytelling is a whole other conversation, check out this article to learn more about public speaking and how storytelling factors into it: Public Speech Into Story: 3 Steps To Telling A Captivating Story

Here the pictures are the heroes, and while words are important, make them complementary to what you are speaking.
Starting off with a joke is also a very popular trick and I think why should it be this or that, why should it be a joke or a story, why can’t it be a humorous story?
Now don’t go fretting about because it doesn’t have to be fictional, it could even be an anecdote from your experiences or maybe one comic strip you found online.
When it comes to humorous speeches, it can be quite intimidating, but here is an article I think will help you wade through these waters: A Guide To Using Humour In Your Speech
Idea 8: Examples
This is a great way to introduce your topic to a crowd that doesn’t know your topic well. Create examples or situations to help your audience gain a smooth entry into your presentation.
It is like math, it is fun when you understand, and that means you care and give attention to it.
You can also use case studies or make your examples into stories to make it more subtle and seamless.

Here is where a traditional topic, sentence and image layout of an opening slide is best suggested. You can build this in any direction and still be able to relate to your slide.
Idea 9: Hard Facts
Facing facts instances that are always either pleasantly welcomed or hard to swallow. Hitting the audience with hard facts works really well, especially if what you are going to talk about is a difficult or sensitive issue.
An astonishing fact is bound to catch people’s attention and you can always use it to your advantage!
According to Femme International, over the last 20 years, the sanitary pad sector has bloomed and advanced; they have taken over the industry and 85% of menstruating women in the country use napkins. As society progressed and the taboo on periods were lifted from many regions, a new problem came up. One which is really harmful. We all know that the blood that comes out during our periods is harmful and full of bacteria. Now include this bacteria filled blood with a pad which takes 500-800 years to decompose. That’s right, 500-800 years of a used sanitary napkin breeding bacteria in rivers, drains, soil and the sea. A menstruating woman uses 15-20 pads for one cycle. Which sums up to 7,200-9,600 pads over an average period of 40 years. This is just for one woman. According to UNICEF roughly 26% of the world’s population are menstruating women. This means that 2.28 BILLION women are going to use over 9,000 pads EACH during their menstruating years.

Always try to not keep your introductory slides text heavy, but when starting with facts, try to highlight them, notice how the topic and the image are not very prominent but play their part in bringing together the entire slide while the first thing you read is the fact, underlined and set in the middle.
Try to play around with the layouts, figure out what suits your needs the best.
Idea 10: Controversial Statements
Who doesn’t love controversies?
Even if we know something is clickbait, it still catches our eye. Even if we know something to not be possible, when someone says it – with conviction, our ears do perk up.
It doesn’t have to be something extraordinary, just not ordinary enough that it catches people’s attention and in the end, you can always use it to connect your conclusion to your introduction.
Here is a great TEDTalk that would help you understand what I am talking about.
If you plan to use this method, it is easier to dive into your slides after you’ve made the statement and start elaborating on it instead of right at the beginning, it could start with your topic or some proof or where ever your presentation takes you!
Final Thoughts
A presentation carries as much personality as its maker, if you want the right impact you need to use the templates, infographics and tools available to you to the fullest, but remember, there is a thing called “too much” as well.
The easiest way to kill it with your presentations is to keep it neat, in your aesthetic and to the point. Make it engaging, make it colourful, make it black and white. It would work perfectly if it bounces off your personality on stage.

Enroll in our transformative 1:1 Coaching Program
Schedule a call with our expert communication coach to know if this program would be the right fit for you

High-Stakes Presentations: Strategies for Engaging and Influencing Senior Leaders

Crisis Leadership 101: Cultivating Empathy While Exercising Authority

Lost Voice? Here’s How to Recover Sore Throat and Speak Again
- [email protected]
- +91 98203 57888
Get our latest tips and tricks in your inbox always
Copyright © 2023 Frantically Speaking All rights reserved

6 Examples of Killer Speech Openings That You Can Use
Think about a speech you saw or heard that stuck with you. What is it about that speech that makes you remember it long after you’ve heard it? The speaker was probably dynamic and engaging or the topic was compelling and of particular interest to you. The speech probably had a killer opening as well.
The introduction to a speech is one of its most crucial aspects. It’s where you grab your audience’s attention and set the tone for the rest of the speech. If your speech opening isn’t memorable, the rest of your speech probably won’t be either.
To help you understand the impact a good opening can have, here’s a list of six killer speech intros. The fact that these speeches are so memorable shows why public speaking is important and why your introduction should be strong.
Julian Treasure: Start with a question
“The human voice: It’s the instrument we all play. It’s the most powerful sound in the world, probably. It’s the only one that can start a war or say ‘I love you.’ And yet many people have the experience that when they speak, people don’t listen to them. And why is that? How can we speak powerfully to make change in the world?”
Julian Treasure’s speech “How to speak so that people want to listen” showcases a tried and true speech opening technique: asking a question. You shouldn’t ask just any question, though. It should be a compelling, thought-provoking question that doesn’t have a straightforward answer.
Asking the audience what they had for lunch that day isn’t compelling, unless you have an interesting reason for asking it that you follow up with. Treasure’s compelling question doesn’t require an immediate answer but instead asks you to think.
Ric Elias: Incredible story
“Imagine a big explosion as you climb through 3,000 ft. Imagine a plane full of smoke. Imagine an engine going clack, clack, clack, clack, clack, clack, clack. It sounds scary. Well, I had a unique seat that day. I was sitting in 1D.”
One way to grab the attention of your audience right away is to tell an exciting or unbelievable story. Ric Elias’s speech about surviving a plane crash goes straight into the action, asking the listener to imagine unbelievable situation he was in. With an opening like that, who wouldn’t want to keep listening to find out what happens next?
To make your intro story more effective, don’t finish it right away. Come back to it later in the speech so that your audience is held in anticipation.
Dan Pink: Make a “confession”
“I need to make a confession at the outset here. A little over 20 years ago, I did something that I regret, something that I’m not particularly proud of.”
Dan Pink’s speech grabs your attention right away because he makes a confession. He’s letting you in on a secret about himself or something he did and you want to listen because you want to hear it.
Letting your audience know a secret or confessing information about yourself makes them feel exclusive, like they have access to something that others don’t. Exclusivity always piques interest.

Jane McGonigal: Provocative statement
“I’m a gamer, so I like to have goals. I like special missions and secret objectives. So here’s my special mission for this talk: I’m going to try to increase the life span of every single person in this room by seven and a half minutes. Literally, you will live seven and a half minutes longer than you would have otherwise, just because you watched this talk.”
Starting a speech with a provocative statement is similar to starting with a question. You make the audience think. You surprise them or say something unexpected and they keep listening to hear an explanation.
Jane McGonigal uses this tactic in her speech , claiming she will expand the life span of every audience member by seven and half minutes. It’s intriguing, provocative, and makes you wonder instantly how she’s going to do it.
Pamela Meyer: Set up a problem (then solve it)
“Okay, now I don’t want to alarm anybody in this room, but it’s just come to my attention that the person to your right is a liar. Also, the person to your left is a liar. Also, the person sitting in your very seats is a liar. We’re all liars.”
People love hearing about a big problem and then knowing the solution — it’s a common framework from literature that you can borrow for your speech. By telling her entire audience that they are liars, Pamela Meyer sets up a problem that seems difficult to solve: how to know who is a liar. She unravels the solution in the rest of her speech , explaining how to become a liespotter.
Sir Ken Robinson: Humor
“Good morning. How are you?” (Audience replies “Good”) “It’s been great, hasn’t it? I’ve been blown away by the whole thing. In fact, I’m leaving.”
Humor can be an impactful way to start a speech, but it’s also risky. Your joke might fall flat and leave everyone feeling uncomfortable. If you tell the right joke, however, it can go over well.
Sir Ken Robinson’s speech , “Do schools kill creativity?” is one of the most watched TED Talks of all time and for good reason. His speech is engaging and dynamic, centering on a topic that almost everyone can easily relate to. What could’ve been a boring start — “Good morning. How are you?” — becomes a funny and memorable speech opening thanks to Robinson’s timing and deadpan delivery.
Make your speech opening memorable
If you want to deliver an engaging address that your audience members will speak about years later, be sure to craft an unforgettable opening. By using any of the above techniques, you’re likely to have a more solid start to your speech. Who knows? Maybe your speech opening will be so good it’ll end up on a list just like this one.
You May Also Like

The Secret to Good Storytelling

12 Pretty Good Hooks for Your Next Presentation
“the next three minutes will determine whether you succeed or fail on your next presentation.”.
Every presentation has three main parts – the beginning, the middle, and the end. And among these, the beginning is easily the most important.
You have about 30 seconds to get the audience interested in what you have to say. If that doesn’t happen, anything else you do afterwards will soon be forgotten.
To do that it helps to have a “hook” to grab their attention. Here are 12 tried and true ways to hook your audience and get your presentation off on the right foot.

Why You Need to be Captain Hook
At the beginning of every presentation, there are two main things going on.
- You are trying to introduce your topic and convince people why they need to hear it.
- Your audience is trying to decide whether this will be worth their time.
A good hook is one that quickly gets everyone oriented and engaged, introduces your topic, and makes them willing to listen to more of what you have to say.
A good hook orients, engages, and makes the audience interested in hearing more. Click To Tweet
There are lots of effective ways to do this. The main thing is to do something interesting.
Trying to hook an audience? The main thing is: Be Interesting! Click To Tweet
It’s hard to think of a quicker way to shut down the audience than to begin by mumbling,
“Hello, my name is Bob Smith, and for the next 45 minutes I’m going to be talking about the importance of socks.”
I don’t know why Bob’s talking about socks, and I don’t know why I should listen. I’m already lacing up my track shoes and getting ready to sprint for the door.
So if it falls to you to give the next presentation on the importance of socks (or any other topic), here are 12 ways you can use to engage the audience from the very beginning. I’ll continue with the sock example to help out poor Bob.
Note: If you would like a one-page condensed version of these 12 hooks, plus presentation tips to help them work for you, hit this button:
12 Pretty Good Hooks About Socks
1. Make an Outrageous or Provocative Claim . It’s the same way I started this post. Hopefully it caught your attention and made you want to read more. The same goes for presentations. Be bold, original, challenge common beliefs, then back up your claim.
“Socks are more important than food.” That’s what a man named Kiwi said recently. Kiwi was a homeless man living on the streets of Toronto, Canada. He said that he knew of many pantries and shelters where he could get food, but there was no way he could walk the streets of his city without socks. Socks meant everything to this man and many like him. And if you think about it, socks are very important to all of us…
(Got this quote from The Joy of Sox , a charitable organization that donates socks to the homeless.)
2. Tell a Joke. This is a classic technique that can work well as a way to relax and make the audience receptive. It’s a good idea to try the joke out on some friends first, to make sure they think it’s funny too. Keep it clean, and be ready to move on if you don’t get the laughter you thought you would.
On the first day of summer camp, the director informed all the little campers that he expected them to put on a fresh pair of socks every day. Two weeks later, Johnny failed to appear at the morning flag raising. The director found him still in his cot.
“Why aren’t you lined up with the other boys?” He asked.
“I can’t get my shoes on over fourteen pairs of socks!”
Clearly the boy knew that it was important to have his socks, but didn’t understand why or how the socks were important…
This site at ajokeaday is clean and has a long list of joke categories and a pretty good search function to help you find just the right joke to get started.
3. Tell a Story. Stories remind your audience that you are human, make you relatable, and take down protective barriers people will erect when they think people will be talking at them. Start immediately with the story. You can back up and introduce yourself later.
“ Change your socks ” the instructor ordered. We had just forded a river in the middle of the night and our feet were soaking wet. We were carrying three pairs of socks on this mission. The first was already dirty, and now the second was soaked. But we didn’t want to change our socks. Because in two miles we were supposed to cross another river and we would get wet again. But the instructor didn’t want to hear that. He wanted to make sure we knew he was in charge. Presenting him with the facts only appeared like a challenge to his authority, so he simply said slowly and more deliberately, “ Change. Your. Socks .”
So we changed into our last dry socks by moonlight, he checked every one of us, then we marched on. And we crossed that next river and got wet. Our feet were soaked and sore for the next two days. For a while, it was all I could think about – the foolish order to change our socks. It was the maddest I had ever been. Because when it comes right down to it, the simplest things are the most important in life. The trust of a loved one; food; and dry socks. Most of us take socks for granted…
The best stories are personal ones from your own experience, but you can also find good ones on the internet. This page at businessballs.com indexes their stories by title and topic.
4. Show a Video . Let someone else break the ice for you with a video. It’s an unexpected way to begin, so people will be instantly interested, plus people just like to watch video. Keep it brief and related to the topic in some way. It can be a great way to show the audience what you are talking about, not just trying to describe it. Absolutely double check and rehearse to make sure the video works wherever you will be presenting, and be prepared to react if it fails anyway.
So, who knew you could make a smart phone holder out of an old sock? Well in fact there are lots of good uses for socks, and some actually involve putting your feet into them. Socks are one of the most underrated items of clothing…
5. Ask a Couple Questions. Some people will want to answer, but even if they don’t do it out loud, they will be considering the answer in their minds, so they will be engaged. Asking several questions in a row will stretch the audience’s mind further while also serving as an introduction to your topic. Be sure to pause after each question for best effect.
What do you think is the most important piece of clothing you own? What makes it important? Is it the protection it provides? How often you wear it? The warmth it offers? The way it makes you look? How it makes you feel? For me, there is nothing more important that having on a nice clean pair of dry socks…
6. Show a Picture . This can be a variation of the story method. Share a picture of a person and talk about him, or of a place and why it is important, or of something else related to your subject. People will look at the picture instead of you, so it takes a little pressure off. And the picture itself serves to introduce your topic. Half the job is done before you even open your mouth.

7. Ask a Rhetorical Question . It’s a question that can’t necessarily be answered, but it gets people thinking, and helps you point them in the direction you want to go.
Are socks what truly define us as human beings? Are socks what differentiate us from all the other forms of life on the planet? After all, nobody else on earth is wearing them except us. Can they be that important? These are clearly questions for the ages, and well worth our consideration…
8. Set an Expectation . Not only is it a good idea to let people know what you will be talking about, you will engage their attention much better if you give them something interesting to look forward to.
By the end of this presentation, not only will you know how important socks are in our lives, you will have ten fun facts about socks that you probably didn’t know, and four new sock jokes that you can use to amaze and amuse your friends.
9. Show Them an Object. Bring something with you that you can hold up and talk about. This is a good way to quickly capture the attention of the audience and introduce your topic at the same time.

10. Reference an Historical Event . If the day, week, month, or year is unique in any way that you can relate to your topic, that can be another way to make the topic seem more real or relevant.
Did you know that it was only 135 years ago, on a day very like today, that John Nelson, a Swedish immigrant to the United States, patented the sock-knitting machine? And that one invention was enough to not only change the history of footwear, but lead to the development of a child’s toy as well. Socks have had a huge impact on us…
At this New York Times link , you can see what happened today in history, or pick any other date and see what was going on.
11. Use a Quote . Find a quote from someone recognizable, then think about how you can tie it into your presentation, or turn it on its head.

Over at Brainyquote.com you can find lots to choose from and can even search by topic. Another good source is Wisdomquotes for lots of good quotes by topic or author.
A variation on this idea would be to make the quote your first slide – nothing but the quote in giant words; possibly with a picture of the person who said it.
12. Ask, “What if…?” or “Imagine…” Ask them about something that changes their perspective, like what if you could fly, read minds, be debt-free, go backward or forward in time. You can adapt this to nearly any presentation and it will immediately cause your audience to engage their minds. Ask the question, pause, ask it again for best effect.
Imagine a world without socks. (pause) Imagine… a world without socks. What would it be like? What would change? How would your life be different? (pause) When you think about it, socks are critically important….
These are just 12 possible ways to hook your audience, and there are infinite variations on these ideas. You can even combine them – tell a funny story while showing a picture or turning a quote on its head. The keys are to keep it interesting, original, and fairly brief. Remember, you only have about 30 seconds before they decide whether or not they want to keep listening. Use that time well, and you will be on your way to making a memorable presentation.
If I was able to help Bob get you even the least bit interested in something as mundane as socks, think of what you can do with a far more interesting topic!
With that, since we’re on a sock “thing” today, I’ll leave you with an ode to socks I came across by Chilean poet and diplomat Pablo Neruda . Maybe if I had quoted this to my instructor so long ago, he’d have let me save my last pair of dry socks for after the final river crossing…
Ode to my Socks
Mara Mori brought me a pair of socks which she knitted herself with her sheepherder’s hands, two socks as soft as rabbits. I slipped my feet into them as if they were two cases knitted with threads of twilight and goatskin, Violent socks, my feet were two fish made of wool, two long sharks sea blue, shot through by one golden thread, two immense blackbirds, two cannons, my feet were honored in this way by these heavenly socks. They were so handsome for the first time my feet seemed to me unacceptable like two decrepit firemen, firemen unworthy of that woven fire, of those glowing socks.
Nevertheless, I resisted the sharp temptation to save them somewhere as schoolboys keep fireflies, as learned men collect sacred texts, I resisted the mad impulse to put them in a golden cage and each day give them birdseed and pieces of pink melon. Like explorers in the jungle who hand over the very rare green deer to the spit and eat it with remorse, I stretched out my feet and pulled on the magnificent socks and then my shoes.
The moral of my ode is this: beauty is twice beauty and what is good is doubly good when it is a matter of two socks made of wool in winter.
Wool socks in winter – amen to that!
Egyptian Socks photo; no changes made – https://commons.wikimedia.org/wiki/File:Early_socks.jpg
Sock joke – source: http://www.jokebuddha.com/Socks/recent/4#ixzz3fJpmAwQ7
The Joy of Sox charitable organization donates socks to the homeless http://www.pointsoflight.org/blog/2014/06/04/when-socks-are-more-important-food%E2%80%99
Sock Monkey photo; no changes made – https://www.flickr.com/photos/poplinre/2153204704/ –

Ken served for 26 years in the Infantry, retiring as a Colonel. From leading patrols in the Korean DMZ, to parachuting into the jungles of Panama, to commanding a remote outpost on the Iran-Iraq border, he has learned a lot about leadership, and has a passion for sharing that knowledge with others. Look for his weekly posts, check out his online courses , subscribe below, or simply connect , he loves to talk about this stuff.

GET LEADERSHIP TOOLS
RAPIDSTART LEADERSHIP © 2015-2023 Ι ALL RIGHTS RESERVED

There's a lot here
Would a one-page cheat sheet help, (no strings attached), they are ready to follow..., ...are you ready to lead, subscribe now.
Killer Presentation Key #3: Rock the Introduction

The third key to creating a killer presentation is to rock the introduction . Now, there will be situations where you will introduce yourself and in other circumstances, other people will introduce you. But here is some valuable advice: If other people are introducing you, do not let them introduce you with their own words. Give them the words you would like them to use when they introduce you.
It may be that they do not know you and therefore may not present you in the best light, so help them out. You will likely find that the people introducing you at events or the people who invited you to speak in the first place will be grateful that you gave them material for the introduction, because they are probably also working from notes. They might even be panicking about needing to stand up in front of everyone to introduce you. You are worried about your thirty-minute presentation and here they are worried about a thirty-second introduction. You have it in your power to help them and be there for them. Create a brief introduction by including a little about your background and anything else you need them to know.
Earn The Right
You will also need to introduce yourself as well once you stand up to speak but the main point is you have to earn the right. After all the work that has gone into this program, the people sitting there need to believe that you are the expert and they will only believe this if you tell them. You’re going to accomplish this by telling a short story about your life. What have you done with your life? You could take a humorous approach and tell them a story about one of the dumbest things you have ever done in your life so that they don’t have to repeat your mistake. It doesn’t need to be about a great accomplishment. It could be a silly story, but the key is it needs to contain emotion.
This is called an ETR. Earn The Right. You are earning the right- the right to be standing in front of them and teaching them, so you have to present your credentials. The objective is to let people know through ETR is who you have helped, how many, and perhaps for how long you have been helping people. How have you assisted them and how were their lives changed based on the information you shared with them?
Qualify Yourself and Prime Your Audience
After you have qualified yourself with this information, you also want to prime the audience by letting them know what they are going to get out of this presentation. You want to communicate to them, “Hey listen, by the end of tonight, my intention is that you are going to walk out of here with strategies that will change your life in whatever your field is.”
So, let them know that by the end of your workshop- however long it will be- how they will be changed after being with you. It gives them a structure, a framework, on which to hang their own expectations and to get the most out of your presentation. What problem are you going to solve for them, and what will they come away with after investing their time in listening to you? Priming your audience will increase their confidence in you and you will be able to feed off their confidence to give an influential, memorable presentation.

Inspirationfeed
Inspiring and educating bright minds.
7 Steps to Giving a Killer PowerPoint Presentation

Last Updated on February 26, 2024
Table of Contents
Widely accepted as the most useful and accessible way to create visual aids ready to share with an audience, PowerPoint presentations are often poorly constructed making them boring and arduous to sit through.
With so many uses and tools to help you give a fantastic presentation every time, it’s frustrating to see so many bad examples. Some sources claim that up to 50% of presentations are ineffective.
A well-designed slideshow allows the presenter to maintain eye contact with the audience, creating an engaging experience for all involved. On the flip side, garish colour schemes, incorrect font sizes and poor image selection can turn your points from being clear to confusing. Here are our 7 best training tips for a better presentation .
1. Get Organized with Slide Sections
Ok, so this one is obvious but you’ll be surprised how many people dive straight into creating a presentation without setting a plan and laying out the groundwork.
Ask yourself what the key messages are that you want to get across to your audience. Whether it’s updating the team on sales revenue figures or you’re pitching a sale to a potential client; be clear about your objectives and stick to the point.
PowerPoint offers some fantastic organization tools for users of all levels to utilize. Most notably Slide Sections , which enable simpler collaboration for teams and better organization for individuals. Similar to putting files into a folder, you can arrange slides into sections. Create new headers, drag and drop slides all without affecting the layout of your presentation.
2. Create a Theme with Slide Master
The best slideshow presentations carry a theme throughout. In their simplest form, standardising fonts and well-designed themes are a great starting point.
PowerPoint of course offers standard themes for you to choose from, along with recommending fonts that go well together – for those times when more than one font helps to liven up your slides.
3. Use Easy to Read Fonts with Suggested Fonts
There’s little more frustrating that your audience all facing the projector squinting – perhaps if they’re facing the floor snoring. Poorly selected fonts and colour schemes can result in hard to read slides. This detracts away from the your presentation and the points that you’re trying to make.
4. Get straight to the point
Great slideshows are all about storytelling. With a beginning middle and end, your presentation will have a common plot.
Bullet points are a great way of getting to the point. Highlighting key facts and figures give the audience a quick indication of the topic without having to sift through a pile of adjectives first.
5. Use the slides as a guide
We can all remember sitting through a presentation where the presenter has read each slide word for word. Aside from being painfully boring, it’s also a massive waste of time for everybody involved. These slides tend to be word dense and could be better distributed as a white paper or a report.
Using bullets is a great way of breaking down your points into manageable chunks. Glancing at these slides allows you to elaborate or adlib whilst keeping eye contact with the room and in turn engaging with your audience.
6. Be Visual
Sure each slideshow has its own subject and aims, but we all know that pictures tell a thousand words. Used to reinforce your points or to inject a little subtle humour, images help to keep an audience captivated throughout.
As a rule of thumb, it also pays to keep things simple. Ok, so we’ve said it a hundred times already but the faster that your audience gets on the same page and understands your point, the more successful the presentation is likely to be.
Our top tip is to use charts instead of tables, with the former tending to be a graphical analysis of the data represented in the table. If the audience requires additional statistics, these should be included in handouts for later analysis.
7. Presenter View in PowerPoint
If you’re lucky enough to have the latest version installed on your PC, you’ll be able to take full advantage of PowerPoint’s awesome new features.
Presenter View enables you too see what’s coming next before the audience. Helping to prevent embarrassing mistakes and keeping you one step ahead of the presentation is sure to make things run much more smoothly.
Other handy tools include a laser pen that is controlled by your mouse or track-pad and a zoom tool that allows you to focus on a specific area of the slide, great for table and charts.

Posted by: Terence Craven
Terence is a copywriter for Best STL. When he's not blogging, he can often be found at the gym or running through the British countryside.

Communication , Featured
18 tips for killer presentations.

Jerry Seinfeld has a skit where he points out that studies show public speaking is a bigger fear than death. That means, he claims, that if you are going to a funeral you are better off in the casket than doing the eulogy. While there isn’t a lot you can do to melt away your anxiety, a the best start is simply to make a better presentation.
Becoming a competent, rather than just confident, speaker requires a lot of practice. But here are a few things you can consider to start sharpening your presentation skills:
- 10-20-30 Rule – This is a slideshow rule offered by Guy Kawasaki. This rule states that a powerpoint slide should have no more than 10 slides, last no longer than 20 minutes and have no text less than 30 point font. He says it doesn’t matter whether your idea will revolutionize the world, you need to spell out the important nuggets in a few minutes minutes, a couple slides and a several words a slide.
- Be Entertaining – Speeches should be entertaining and informative. I’m not saying you should act like a dancing monkey when giving a serious presentation. But unlike an e-mail or article, people expect some appeal to their emotions. Simply reciting dry facts without any passion or humor will make people less likely to pay attention.
- Slow Down – Nervous and inexperienced speakers tend to talk way to fast. Consciously slow your speech down and add pauses for emphasis.
- Eye Contact – Match eye contact with everyone in the room. I’ve also heard from salespeople that you shouldn’t focus all your attention on the decision maker since secretaries and assistants in the room may hold persuasive sway over their boss.
- 15 Word Summary – Can you summarize your idea in fifteen words? If not, rewrite it and try again. Speaking is an inefficient medium for communicating information, so know what the important fifteen words are so they can be repeated.
- 20-20 Rule – Another suggestion for slideshows. This one says that you should have twenty slides each lasting exactly twenty seconds. The 20-20 Rule forces you to be concise and to keep from boring people.
- Don’t Read – This one is a no brainer, but somehow Powerpoint makes people think they can get away with it. If you don’t know your speech without cues, that doesn’t just make you more distracting. It shows you don’t really understand your message, a huge blow to any confidence the audience has in you.
- Speeches are About Stories – If your presentation is going to be a longer one, explain your points through short stories, quips and anecdotes. Great speakers know how to use a story to create an emotional connection between ideas for the audience.
- Project Your Voice – Nothing is worse than a speaker you can’t hear. Even in the high-tech world of microphones and amplifiers, you need to be heard. Projecting your voice doesn’t mean yelling, rather standing up straight and letting your voice resonate on the air in your lungs rather than in the throat to produce a clearer sound.
- Don’t Plan Gestures – Any gestures you use need to be an extension of your message and any emotions that message conveys. Planned gestures look false because they don’t match your other involuntary body cues. You are better off keeping your hands to your side.
- “That’s a Good Question” – You can use statements like, “that’s a really good question,” or “I’m glad you asked me that,” to buy yourself a few moments to organize your response. Will the other people in the audience know you are using these filler sentences to reorder your thoughts? Probably not. And even if they do, it still makes the presentation more smooth than um’s and ah’s littering your answer.
- Breathe In Not Out – Feeling the urge to use presentation killers like ‘um,’ ‘ah,’ or ‘you know’? Replace those with a pause taking a short breath in. The pause may seem a bit awkward, but the audience will barely notice it.
- Come Early, Really Early – Don’t fumble with powerpoint or hooking up a projector when people are waiting for you to speak. Come early, scope out the room, run through your slideshow and make sure there won’t be any glitches. Preparation can do a lot to remove your speaking anxiety.
- Get Practice – Join Toastmasters and practice your speaking skills regularly in front of an audience. Not only is it a fun time, but it will make you more competent and confident when you need to approach the podium.
- Don’t Apologize – Apologies are only useful if you’ve done something wrong. Don’t use them to excuse incompetence or humble yourself in front of an audience. Don’t apologize for your nervousness or a lack of preparation time. Most audience members can’t detect your anxiety, so don’t draw attention to it.
- Do Apologize if You’re Wrong – One caveat to the above rule is that you should apologize if you are late or shown to be incorrect. You want to seem confident, but don’t be a jerk about it.
- Put Yourself in the Audience – When writing a speech, see it from the audiences perspective. What might they not understand? What might seem boring? Use WIIFM (What’s In It For Me) to guide you.
- Have Fun – Sounds impossible? With a little practice you can inject your passion for a subject into your presentations. Enthusiasm is contagious.
What tips do you have for making killer presentations?

How to Use a Planner Effectively

How to Be a Better Planner: Avoid the Planning Fallacy

5 Best Apps to Help You Delegate Tasks Easily

Delegating Leadership Style: What Is It & When To Use It?

The Fear of Delegating Work To Others

Why Is Delegation Important in Leadership?

7 Best Tools for Prioritizing Work

How to Deal with Competing Priorities Effectively

What Is the RICE Prioritization Model And How Does It Work?

4 Exercises to Improve Your Focus

What Is Chronic Procrastination and How To Deal with It

How to Snap Out of Procrastination With ADHD

Are Depression And Procrastination Connected?

Procrastination And Laziness: Their Differences & Connections

Bedtime Procrastination: Why You Do It And How To Break It

15 Books on Procrastination To Help You Start Taking Action

Productive Procrastination: Is It Good or Bad?

The Impact of Procrastination on Productivity

How to Cope With Anxiety-Induced Procrastination

How to Break the Perfectionism-Procrastination Loop

15 Work-Life Balance Books to Help You Take Control of Life

Work Life Balance for Women: What It Means & How to Find It

6 Essential Mindsets For Continuous Career Growth

How to Discover Your Next Career Move Amid the Great Resignation

The Key to Creating a Vibrant (And Magical Life) by Lee Cockerell

9 Tips on How To Disconnect From Work And Stay Present

Work-Life Integration vs Work-Life Balance: Is One Better Than the Other?

How To Practice Self-Advocacy in the Workplace (Go-to Guide)

How to Boost Your Focus And Attention Span

What Are Distractions in a Nutshell?

What Is Procrastination And How To End It

Prioritization — Using Your Time & Energy Effectively

Delegation — Leveraging Your Time & Resources

Your Guide to Effective Planning & Scheduling

The Ultimate Guide to Achieving Goals

How to Find Lasting Motivation

Complete Guide to Getting Back Your Energy

How to Have a Good Life Balance
Explore the time flow system.

About the Time Flow System

Key Philosophy I: Fluid Progress, Like Water

Key Philosophy II: Pragmatic Priorities

Key Philosophy III: Sustainable Momentum

Key Philosophy IV: Three Goal Focus

How the Time Flow System Works
How to Create Killer Presentation Slides (Key Components and Tips)
Last updated on April 16th, 2024

Powerful slide presentations are visually appealing, not visually distracting. They effectively convey a specific message to a certain group of people for various purposes. However, not everyone knows about the crucial components of a killer presentation that can impact the listeners’ minds.
Slide shows are easy to produce, update, and present if you know how to grab the attention of your audience . Creating powerful presentations is one of the skills that will let you differentiate from many professionals, business leaders, public speakers as well and students. That is why you must be aware of specific components of engaging presentations with tips to present in front of any listener. So, how to make a good slideshow presentation? In this article, we present tips that will let you make killer presentations that stand out from the crowd.
Why putting in the effort to create killer presentations?
In the modern age of technological revolution, digital means of communication have prevailed in almost every type of industry. If you want to deliver an idea, a message, a business proposal, or an argument, slide decks are one of the most effective tools to achieve these objectives.
1. Presentations are powerful sales tools
PowerPoint presentations are effective sales tools as they are used for selling ideas, products, and services in almost all types of industries and for a variety of business models, B2B sales, B2C, social selling, etc. A speaker or presenter may have his own method of delivering a message but the visual appeal of the sales deck proves to be very helpful in resonating with the minds, thoughts, and ideas of the listeners.

By using sales presentations or sales decks, a salesman can inform his prospective clients by informing them of the potential benefits of his products or services. He can then convince them by identifying their problems and offering his product or service as a solution. In the end, with the aid of a structured sales presentation that includes compelling graphics, diagrams, infographics, and visual slides, he can persuade his listeners to purchase his product or service. A salesman with a top-notch slide presentation can convert his listeners into customers.
2. Presentations are important in public speaking
Being a good orator or public speaker is not enough to leave a long-lasting impact on your audience during your public speaking activities. You will need the assistance of other techniques and tools as well to become a powerful public speaker . That is why presentations are very useful in the public speaking process as they facilitate the speaker in conveying his message through the visual aid.
According to analysts, a piece of visual information gets perceived and understood 60,000 times faster than textual information . So, the recipe to deliver a top-notch public speech is to have a well-organized and audience-focused speech along with organized and structured visuals and graphics in the form of a presentation.
You don’t have to be a graphic designer or Adobe Photoshop expert to create and design a killer presentation for your next event or activity. You can take advantage of free templates and presentation slides provided by sites like FPPT, where you can download thousands of premade PowerPoint Templates for any topic. In this way, you can focus more on your presentation delivery part than on slide designing and preparation stages.
3. Presentations are essential for promoting brands
To outshine your competition in the ever-vibrant markets, you need to be aware of modern and appealing business tactics and techniques to grab the attention of prospective buyers and prospects.
When business leaders and professionals attend various seminars, exhibitions, or virtual video conferences, slide decks become much more effective tools to promote your services, products, and ultimately brands.
Presentations can be regarded as the language of business communication processes. Therefore, mastering the art of making and delivering efficient slide decks has become more relevant and necessary in the modern business landscape. Putting all the effort into making a killer presentation will help you to differentiate yourself from your competitors.
So, how to give a killer presentation? In the next chapters we will explain how to prepare a great presentation and give it to the audience.
What are the Key Components of a Killer PowerPoint Presentation?
In order to develop and present powerful PowerPoint presentations, you need to be aware of a structured approach to organizing your data and information in the form of slides. Here are some actionable key components of a killer presentation that can help you in achieving your desired goals and help you make your presentation look better.
1. Cover Slide
A cover slide is used to set the tone of the whole presentation topic. Microsoft PowerPoint allows you to create your slides cohesively by offering some basic preset themes and color variations. However, to grab the attention of your listeners, you will need to have a more professionally designed slide presentation. Our website comes in handy in offering a variety of presentation cover PPT slides and designs for almost any kind of presentation topic that you may have. For example, if you want to deliver a presentation on the topic of market trends, you can have a look at the Free Market Trend Analysis PowerPoint Template , to have an idea of how the cover slide will look in this case. You can customize your presentation cover page and slides as per the guidelines of your company’s brand identity or you can choose your own visually appealing graphics to seem more interesting.
2. Agenda or Table of Contents Slide
After designing and setting your cover slide, it is time to provide an overview to your audience about the topic and purpose of your presentation. This is generally done with the help of agenda slides in your presentation.
For example, if you deliver a presentation about a potential product to prospective buyers, you can mention the following upcoming slides in your agenda slide in a numbered manner.
- Who are you?
- What is the problem being faced by users?
- How can you (your company or business) solve this problem?
- Why choose your solution?
- Drawbacks of using other products and the benefits of using your product?
- Questions and Answers Session
Typically, each point in the agenda slide will correspond to a specific slide in your presentation.

In some interactive presentations, designers can place a link so clicking on each of the agenda items will navigate to the desired section and then get back to the agenda. If you want to consider a ToC instead of Agenda, you can learn more on how to create a Table of Contents for your PowerPoint presentation .
3. Storytelling Approach as a Crucial Component
No matter what type of topic is going to be presented, you should always remember that you are delivering the presentations to fellow human beings who have certain desires, feelings, and emotions. In order to leave a profound impact on your audience, your message should resonate with the feelings, and emotions of listeners. That is why, by using a storytelling approach , you can master the art of persuasion.
You need to tell the idea and story behind your product or service and relate it to the needs, wants, and desires of the prospects. The goal is to make your listeners realize that you are exactly a match for their problems and worries. You need to be aware of your buyer personas to become a more effective presenter.
4. Explaining the Problem under Discussion
You need to be aware of the background of your audience to understand their needs, desires, and problems. At this stage, you must mention the problem under discussion in your slide to demonstrate that you have a clear knowledge of the issues being faced by the target customers. Typically you will use a Problem Slide here.
Explain how the problem affects your customers, other businesses, and, if applicable, the whole world. In this way, you can maintain a solid position on the stage as the listeners will be more attentive to you when you discuss their pain points.
5. Offering Your Solution
Now, when the problem being faced by the users has been thoroughly discussed, it’s time to introduce your product, service, or idea as a solution. You have grabbed the attention of your audience until now and listeners are now eager to learn about a possible solution for their worries. This is done with the help of Solution slides.
You need to be creative at this stage while explaining multiple solutions for the issues being faced by the users. You can use PowerPoint shapes to give a visual touch to your textual information. You can also use relevant charts (bar charts, pie charts, line charts, etc.), graphs, tables, infographics , images, gifs, and videos to make your slides more interesting. You can also use metaphors and analogies to represent ideas and explain the solution.
6. Persuasion with Data and Proof
After presenting your solution, it is time to persuade your listeners to take an interest in your offer. If you have developed some reports, you can present their conclusions to potential clients. You can show the data in the form of charts or graphs as they combine both textual and visual elements for better understanding.
You can also mention statistics (key stats) about why your product, service, or idea is better than other industry players. By providing concrete sets of data, you can make the decision-making process of your potential audience much easier. Typically you’d use the key statistics slide to present this information, unless you want to compare metrics vs. other competitors or products. In this case, you will use comparison slides.
7. Call to Action Slide (or CTA slide)
After presenting your proofs and statistics, it is high time to persuade your listeners to respond to your specific call to action. You need to make them aware of all the options that they have and how they can choose a suitable option as per their requirements and budgetary needs.
The goal is to give a clear call to action to your audience. Do not make anything ambiguous or confusing. Your ideas, solutions, and ultimate CTA must be concise, to the point, and compelling in nature.
Sometimes, this call to action is no more than a Thank you slide with the Contact Information, a last slide with a QR Code to drive the audience to visit your website or call you, or just a summary slide with the important information you want the audience to save in their minds.
8. Questions and Answers
In order to continue the dialogue between you and your listeners, you can add a Questions and Answers slide in your presentation. In this way, the audience will have the opportunity to ask any further questions that they may have about your ideas, arguments, offers, or solutions. The goal of having a Q&A session or using Q&A slides in your presentation is to make the communication between the speaker and the audience effective to meet the desired objectives of the overall presentation.
9. Reviewing Your Slide Presentation
At this stage, you should review your entire presentation from the beginning to the end to identify any potential grammatical mistakes and errors in your slides. Make sure that all of your slides are coherent and cohesive with respect to each other and present the topic in a seamless flow.
Ensure all animations, transitions, and visual elements are working and set properly. You can show your presentation to some other professional as well to ask for his opinion. Test the running of the slides in the presentation mode by playing a PowerPoint slideshow and make sure that all points in your slides make sense and seem appealing. Playing the slide show is especially useful if you are applying animations and transitions, or if you want to see how the presentation flows from the first slide to the last slide.
3 Tips for Delivering Powerful Presentations by Defining a Killer Structure
To outshine as a persuasive presenter, here are three useful tips for structuring and delivering your presentations. Here are three effective tips on how to give a killer presentation:
1. Don’t Rely on Pre-set Elements in Your Slides
Every presentation topic has specific customized needs. To fulfill these needs and requirements, the presenter must keep in mind the actual theme of the presentation, the flow of the upcoming topics, and ultimately the desired call to action.
Even when you use premade templates , you can customize specific points, visual slides, fonts, animations, and transitions in your slides. Your PowerPoint Presentation’s theme must complement your needs. Decide whether using bullet points for your topics is a good idea or not. Bullet points are important but they are not always appropriate for all topics. Professional designers may argue that bullet points are boring, and not recommend using them but instead use one of the possible alternatives to traditional bullet points , like block lists, sections or even icons.
If there is no need for animations or transitions in specific slides, don’t use them. Make sure your presentation is not underwhelming for listeners.
2. Be Aware of the Presentation Tools and Equipment
A presenter must be aware of the usage of the tools and equipment necessary for delivering presentations. You should test all the tools before the actual presentation like the projector, HDMI or VGA Adapter, Presentation Remote, Wireless Keyboard, Microphone, Speakers, Smartpen, and presentation devices like Laptops or PCs.
If any malfunctioning or mishap happens during your actual presentation, you may lose the interest, and attention of your audience and ultimately the purpose of your presentation. That is why, it is necessary to be aware of all possibilities related to your presentation equipment and you must have some backup solution in case any unwanted activity happens.
The format of your presentation is also important. While you can play any .pptx or .ppt file in PowerPoint, when you present in a different environment, it would be recommended to save it in a different format (e.g. a PDF presentation ) or host the presentation online.
3. Using Premade Presentation Templates
A smart presentation designer always saves time and focuses on the areas that actually matter for the effectiveness of his presentation delivery mechanism. You don’t need to hire a professional designer for your presentation development or design the slides from scratch all by yourself.
You can save your valued time and resources by downloading professional PowerPoint templates like those offered by SlideModel or the free templates provided by Free PowerPoint Templates totally free of cost. Now, you can focus more on the rehearsal of your presentation than on developing your desired professional pitch deck. Developing and delivering killer presentations was never this convenient before.
Final Words
Presentations are used for a variety of purposes in the business world like selling your products and services, public speaking in seminars and video conferences, and promoting your brands and businesses in exhibitions. Therefore, it is very important for business leaders and professionals to learn about the potential elements of powerful presentations that can win the attention of any audience and how each level can contribute to delivering a presentation, depending on the type of presentation to be created.
A killer slide presentation starts with a solid cover and agenda slides but it also ends with a solid last slide. It involves a storytelling approach to highlighting the pain points of the potential users and offering a solution. It includes slides that contain visually appealing statistical data and information to convince prospective listeners to respond to a specific call to action (CTA). It concludes with a Questions and Answers session in which all of the ambiguities of the listeners are cleared in an effective manner.
By keeping in mind all the above-mentioned tips and key elements related to presentations, any business professional can achieve his desired presentation goals in an actionable and measurable manner.
Leave a Comment Cancel reply
Your email address will not be published. Required fields are marked *
Save my name, email, and website in this browser for the next time I comment.
Sign up to our newsletter
We will send you our curated collections to your email weekly. No spam, promise!
More From Forbes
How to create a killer presentation that will make you a video sensation.
- Share to Facebook
- Share to Twitter
- Share to Linkedin

Making an effective video
Did you know we remember 90% of the video messages we watch, but only remember about 10% of the ones we read? Clearly video is an effective tool and should be an integral part of your marketing strategy. Plus—it can be fun.
From Part 1 of my series, we learned about purpose and branding.
Now let’s talk about creating the presentation itself.
The 5 Parts of a Successful Presentation
Keeping your presentation organized and well-structured will make it easier for you to deliver and your audience to absorb. Here’s a way to do that successfully.
Engage with your audience from the beginning
- Start with a catchy opening. “If you focus on results, you will never change. If you focus on change, you will get results.” This for example, is a quote by Twitter cofounder and CEO Jack Dorsey. It’s catchy, empowering and thought provoking. It grabs the audience’s attention and makes then want to watch more. Try to come up with an opening line, story, joke or piece of advice that will capture the audience’s attention from the get-go.
- Tell them what you’re going to tell them. Look at this like your headline. Tell the audience what you’ll be talking about. This salient point should be the number one message you want the audience to remember when they finish watching. So, make it clear and concise—think of it like a tweet and keep it tight.
- Provide relevant context. Explain the landscape in which you or your product or service is operating. This might include trends that bring rise to a need, changes in lifestyle that create new demand or world events that affect habits or behaviors. Basically, set the stage so listeners understand where your product or service fits into the big picture and why you are the much-needed solution.
- Offer supporting data. Now you need to prove your headline. It can be done effectively using stories, analogies or data. But don’t use too many numbers. The human brain can’t absorb lots of figures at once, especially if it is not physically seeing them written.
- Close with a call to action. Don’t close your video with just a web address or a simple goodbye. By doing that, you miss a huge opportunity. Instead, close with a call to action—to buy, sell, persuade, or even simply engage. Don’t blow it when you’ve come this far.
BlackRock CEO Issues ‘Massive’ Warning After Crypto Flip That Powered A Bitcoin, Ethereum And XRP Price Boom
Ingrid andress’ national anthem at 2024 home run derby draws criticism online, trump—with bandaged ear and misty eyes—appears at republican convention (photos).
Tips For Creating Engaging Content
Think of it this way—the purpose of video content is to provoke, evoke and poke. You want the video to engage with its viewers. Here are a few ways to do that:
Woman talking with not at her audience
Pretend the camera is your best friend. Treat your camera like it’s an audience member and talk with it , not at it. Pretending it’s a familiar face enables you to be more conversational. And that makes you relatable and trustworthy.
Be a storyteller. While data may be retained, stories are retold. And that’s free marketing! So perfect your ability to tell a story—the more surprising and unexpected the better. Stories capture the imagination of your audience and grab their attention by adding emotion and energy to what could otherwise be a plain vanilla presentation. Stories also humanize the storyteller, creating connection and trust between you and your audience.
Add emotion. Content that has an emotional appeal is memorable and impacts the audience in a more profound way. It creates engagement which leads to loyalty.
Create viral content
Create viral content. When appropriate, include humor, happiness, excitement and intrigue to increase the likelihood that your video will be shared and potentially go viral, which makes you a video sensation.
This is Part 2 of my 4-part “Lights...Camera...TAKE ACTION” series on how to effectively use video to grow your business. Stay tuned for my next article on How To Use Your Voice and Hands To Rock Your Presentation.

- Editorial Standards
- Reprints & Permissions
How to Start Your Presentation: 4 Step Formula for a Killer Intro
Video source: lighthouse communications.
Have a presentation coming up? Want to hook you audience from the start? Here is a step by step formula to create interest in your topic, connect to your audience, and establish credibility.
Want to live on the Edge?
About the author.
Screen Name
The Edge for Scholars
We post stuff you shouldn't miss.
May 10, 2018
Join the conversation
Your email address will not be published. Required fields are marked *
You May Also Like

Modernizing Clinical Trial Design & Analysis to Improve Efficiency & Flexibility

Present Less Better: Part 3

Present Less Better: Part 2
Inc. Power Partner Awards Application Deadline Extended: Friday, July 19 Apply Now
- Newsletters
- Best Industries
- Business Plans
- Home-Based Business
- The UPS Store
- Customer Service
- Black in Business
- Your Next Move
- Female Founders
- Best Workplaces
- Company Culture
- Public Speaking
- HR/Benefits
- Productivity
- All the Hats
- Digital Transformation
- Artificial Intelligence
- Bringing Innovation to Market
- Cloud Computing
- Social Media
- Data Detectives
- Exit Interview
- Bootstrapping
- Crowdfunding
- Venture Capital
- Business Models
- Personal Finance
- Founder-Friendly Investors
- Upcoming Events
- Inc. 5000 Vision Conference
- Become a Sponsor
- Cox Business
- Verizon Business
- Branded Content
- Apply Inc. 5000 US
Inc. Premium

5 Tips on Creating a Killer Presentation
Don't make your audience endure one that's dull or mediocre. here's how to reel your listeners in..

How many terrible PowerPoint presentations have you endured during your lifetime? While they may be the standard method of selling , informing, training or otherwise communicating key messages to important audiences, they're easy to screw up. Take some advice from James Ontra, CEO of presentation management platform Shufflrr , who has studied thousands of presentations and knows what goes into creating a stellar one.
1. Create and maintain a slide library.
In other words, why spend time designing content someone in your organization likely already has created? Keep all your presentations in one central place using presentation management software, search for the content you need, update it if you need to and save whatever you've done in the slide library. The trick is to save slides individually with tags under categories, such as "company background," "financials," "services," or "case studies." When saving slides, make sure to include the date they were last updated so that everyone uses the most up-to-date slide.
2. Include video and multi-media content.
There's a reason people love YouTube--video engages people. And if you can get your audience engaged you're halfway to selling your message. What won't hook them? A presentation composed of only bullet points. Just don't overuse video or multimedia content--it can't read a crowd and adjust focus depending on an audience's reaction.
3. Ask your audience questions.
It's another way to get them engaged. Plus, it lets you gauge the audience's interests and comprehension so you can react and tailor your pitch to better reel them in.
4. Pause during remote presentations.
When you're presenting to someone in another location you often can't read their non-verbal cues. Build enough time into your presentation to stop every few minutes to gauge understanding and ask your audience if they have questions.
5. Avoid putting too much content on a slide.
People can read or they can listen, but not both simultaneously. And, they can likely read faster than you can present. If so, not only are they not listening to you, but they are ahead of you. Too much content also can handicap a presenter if he or she ends up reading from a slide. "No one wants to hear you read--they can read it themselves," Ontra says. "Presenters can use speaker notes, but your voice should be used to give context to the content."
Sign up for our weekly roundup on the latest in tech
Privacy Policy
COMMENTS
Frame your story (figure out where to start and where to end). Plan your delivery (decide whether to memorize your speech word for word or develop bullet points and then rehearse it—over and ...
One of the best ways to open your speech with a buzz is to startle or shock them. You can shock an audience in many ways, but they all rest on the major senses of VAKS: Visual. Auditory. Kinesthetic (touch) and Smell. We don't want your audience tasting your talk, but it should leave a good taste in their mouths.
Have a presentation coming up? Want to hook you audience from the start? Then watch this Lighthouse Communications video that gives you a step by step formul...
Introducing yourself in a presentation is pitching yourself to the audience so they stick around for the rest of your talk. Include your background, your unique trait, and who you are while sticking to the context in the first 30-60 seconds of your introduction. Your introduction should be effective and have an interesting hook.
Introducing yourself in a presentation is pitching yourself to the audience so they stick around for the rest of your talk. Include your background, your uni...
What you need - A story, a script and a plan to create a killer presentation. BUT…the process for creating a great presentation should be more like that of making a movie. When you make a movie, filming (the equivalent of creating slides) only happens after you have a story, a script and a plan.
5 Steps to dazzle your audience. https://ruletheroompublicspeaking.com/public-speaking-video-library/Be better by tomorrow. Discover the secrets to giving a ...
The process to create a killer presentation starts six to nine months before the event. That's right! A real killer presentation requires lots of planning, devising, rehearsing, and lots of fine tuning along the way. The actual task of transforming a presentation from muddled to mesmerizing is a matter of hours…spread over a longer period ...
Give rewards for participation. Prompt a discussion in the audience. Build in time for Q&A. Crowdsource questions from the audience. Gather feedback. 1. Plan your storyline. A powerful story can make your whole presentation. Take TED talks, for instance.
Apply the 10-20-30 rule. Apply the 10-20-30 presentation rule and keep it short, sweet and impactful! Stick to ten slides, deliver your presentation within 20 minutes and use a 30-point font to ensure clarity and focus. Less is more, and your audience will thank you for it! 9. Implement the 5-5-5 rule. Simplicity is key.
Let's make the beginning of your presentation count. Here are some ways to start strong: Open with a question or ask the audience a question. Entertain with a great story, prop, or other visual to capture the audience's attention. Use humor, tell a joke, and show vulnerability. Showcase your passion for the topic and/or audience.
Idea 1: Introduction. There is no better way to get the audience to remember you than putting a giant photo of yourself on the screen and going, this is me, - an extremely edited version of me, but still, me. Buddy. No. That was an attempt at being the funny - clever person. Clearly it didn't work.
1. Know who will be in the room.Knowing your audience is the single most important aspect of developing your presentation. Learning about their values, attitudes and beliefs will keep you from ...
1. Research your audience. If you want to influence these people, you need to know what they care about and what motivates them. 2. Include dissenting views. Identify ways your audience may ...
Dan Pink: Make a "confession". "I need to make a confession at the outset here. A little over 20 years ago, I did something that I regret, something that I'm not particularly proud of.". Dan Pink's speech grabs your attention right away because he makes a confession. He's letting you in on a secret about himself or something he ...
Using stories and anecdotes, he highlights what works and what does not. Four key steps will help create a "killer presentation.". First, frame your story. Substance is critical because without it, there is no reason to be presenting. The use of stories and metaphors will also keep the audience engaged. Second, plan your delivery.
How to write a presentation introduction. Consider using the tips below to engage your audience before your next presentation: 1. Tell your audience who you are. Introduce yourself, and then once your audience knows your name, tell them why they should listen to you. Example: "Good morning. My name is Miranda Booker, and I'm here today to ...
12 Pretty Good Hooks About Socks. 1. Make an Outrageous or Provocative Claim . It's the same way I started this post. Hopefully it caught your attention and made you want to read more. The same goes for presentations. Be bold, original, challenge common beliefs, then back up your claim. "Socks are more important than food.".
Killer Presentation Key #3: Rock the Introduction. / Professional Speaking. The third key to creating a killer presentation is to rock the introduction. Now, there will be situations where you will introduce yourself and in other circumstances, other people will introduce you. But here is some valuable advice: If other people are introducing ...
Create new headers, drag and drop slides all without affecting the layout of your presentation. 2. Create a Theme with Slide Master. The best slideshow presentations carry a theme throughout. In their simplest form, standardising fonts and well-designed themes are a great starting point.
Simply reciting dry facts without any passion or humor will make people less likely to pay attention. Slow Down - Nervous and inexperienced speakers tend to talk way to fast. Consciously slow your speech down and add pauses for emphasis. Eye Contact - Match eye contact with everyone in the room.
3 Tips for Delivering Powerful Presentations by Defining a Killer Structure. To outshine as a persuasive presenter, here are three useful tips for structuring and delivering your presentations. Here are three effective tips on how to give a killer presentation: 1. Don't Rely on Pre-set Elements in Your Slides.
getty. Start with a catchy opening. "If you focus on results, you will never change. If you focus on change, you will get results.". This for example, is a quote by Twitter cofounder and CEO ...
About Me. We post stuff you shouldn't miss. The Edge for Scholars. Have a presentation coming up? Want to hook you audience from the start? Here is a step by step formula to create interest in your topic, connect to your audience, and establish credibility.
When you're presenting to someone in another location you often can't read their non-verbal cues. Build enough time into your presentation to stop every few minutes to gauge understanding and ask ...