If you're seeing this message, it means we're having trouble loading external resources on our website.
If you're behind a web filter, please make sure that the domains *.kastatic.org and *.kasandbox.org are unblocked.
To log in and use all the features of Khan Academy, please enable JavaScript in your browser.

AP®︎/College Biology
Course: ap®︎/college biology > unit 7.
- Genetic drift, bottleneck effect, and founder effect
Genetic drift
- DNA spells evolution
- Variation in a species
- Natural selection in populations
- Population genetics
- Genetic drift is a mechanism of evolution in which allele frequencies of a population change over generations due to chance (sampling error).
- Genetic drift occurs in all populations of non-infinite size, but its effects are strongest in small populations.
- Genetic drift may result in the loss of some alleles (including beneficial ones) and the fixation , or rise to 100 % frequency, of other alleles.
- Genetic drift can have major effects when a population is sharply reduced in size by a natural disaster ( bottleneck effect ) or when a small group splits off from the main population to found a colony ( founder effect ).
Introduction
What is genetic drift, genetic drift example, population size matters, allele benefit or harm doesn't matter, the bottleneck effect, the founder effect, attribution:.
- " Population size and genetic drift ," by Douglas Wilkin and Barbara Akre, CK-12 Foundation, CC BY-NC 3.0 .
- " Population genetics ," by OpenStax College, Biology, CC BY 4.0 .
Works cited:
- Krempels, Dana. (2006). Why spay or neuter my rabbit? In Houserabbit adoption, rescue, and education . Retrieved from http://www.bio.miami.edu/hare/scary.html .
- Haw, J. (2013, May 24). Northern elephant seals: Increasing population, decreasing biodiversity. In Scientific american . Retrieved from http://blogs.scientificamerican.com/expeditions/northern-elephant-seals-increasing-population-decreasing-biodiversity/ .
- Genetic drift and the founder effect. (2001). In Evolution . Retrieved from http://www.pbs.org/wgbh/evolution/library/06/3/l_063_03.html .
Additional references:
Want to join the conversation.
- Upvote Button navigates to signup page
- Downvote Button navigates to signup page
- Flag Button navigates to signup page

- Skip to primary navigation
- Skip to main content
- Skip to primary sidebar
- Skip to footer
- Image & Use Policy
- Translations
UC MUSEUM OF PALEONTOLOGY
Understanding Evolution
Your one-stop source for information on evolution
- ES en Español
Genetic drift
Genetic drift is one of the basic mechanisms of evolution.
In each generation, some individuals may, just by chance, leave behind a few more descendants (and genes, of course!) than other individuals. The genes and other genetic elements of the next generation will be those of the “lucky” individuals, not necessarily the healthier or “better” individuals. That, in a nutshell, is genetic drift. It happens to ALL populations — there’s no avoiding the vagaries of chance.

Earlier we used this hypothetical cartoon. Genetic drift affects the genetic makeup of the population, but unlike natural selection , through an entirely random process. So although genetic drift is a mechanism of evolution, it doesn’t work to produce adaptations .
- More Details
- Teaching Resources
Read more about:
- The mechanism of genetic drift, sampling error
- The effects of genetic drift
- Two special cases of genetic drift, bottlenecks and founder effects
- What the neutral theory has to say about the roles of drift and selection in evolution
Teach your students about genetic drift:
- The natural selection game , a classroom activity for grades 9-12.
- Problem-based discussion: Simulations of genetic drift , a set of slides for the college level (PowerPoint download).
Reviewed and updated June, 2020.
Development
Natural Selection
Subscribe to our newsletter
- Teaching resource database
- Correcting misconceptions
- Conceptual framework and NGSS alignment
- Image and use policy
- Evo in the News
- The Tree Room
- Browse learning resources
- Why Does Water Expand When It Freezes
- Gold Foil Experiment
- Faraday Cage
- Oil Drop Experiment
- Magnetic Monopole
- Why Do Fireflies Light Up
- Types of Blood Cells With Their Structure, and Functions
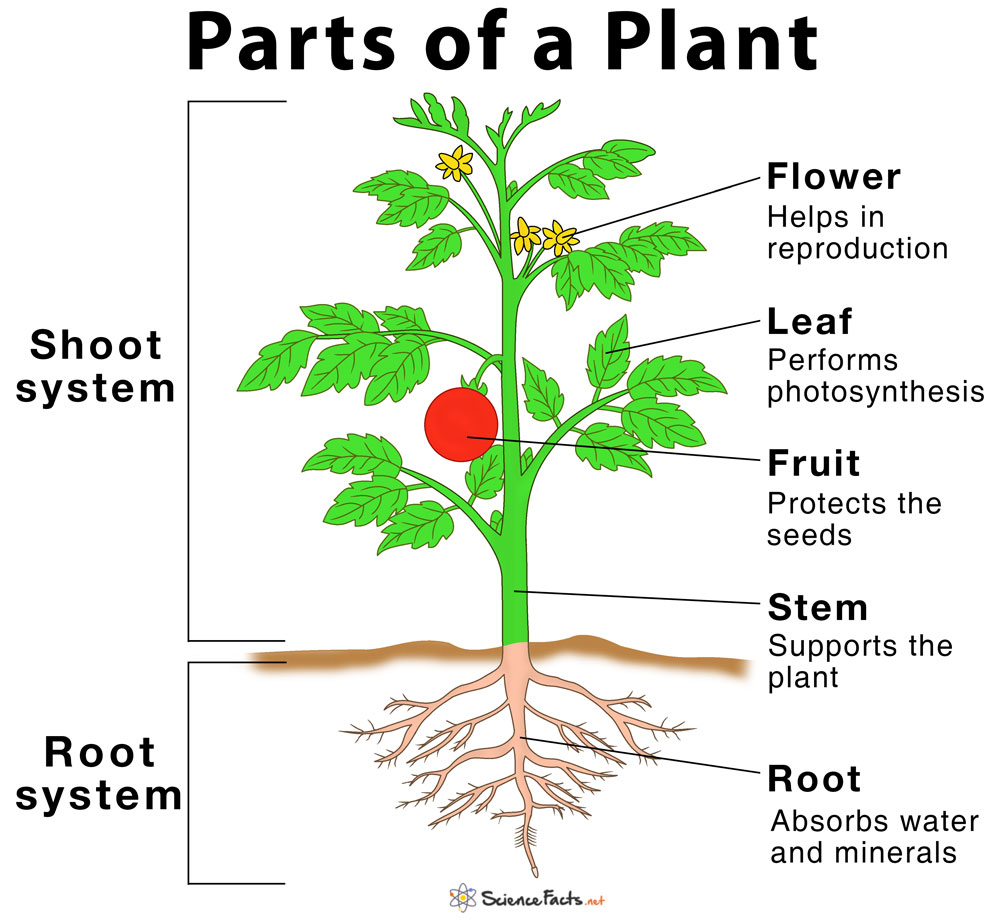
- The Main Parts of a Plant With Their Functions
- Parts of a Flower With Their Structure and Functions
- Parts of a Leaf With Their Structure and Functions
- Why Does Ice Float on Water
- Why Does Oil Float on Water
- How Do Clouds Form
- What Causes Lightning
- How are Diamonds Made
- Types of Meteorites
- Types of Volcanoes
- Types of Rocks
Genetic Drift
Genetic drift, also known as allelic drift or the Wright effect, refers to the random change in the allele frequency of a particular gene in a population. It is one of the basic mechanisms of evolution with natural selection . Unlike the latter, it does not depend on an allele’s beneficial or harmful effects. It can majorly influence small populations where chance events have a more significant impact.
Although random, genetic drift can have long-lasting effects on the genetic makeup of a population. The new population can be genetically distinct from the original group, which led to the hypothesis that genetic drift may form a new species. However, it does not bring adaptation to a population.

Types of Genetic Drift
Genetic drift can be of two types: 1) Bottleneck Effect and 2) Founder Effect. They differ by the kind of event that causes them.
1. Bottleneck Effect
It occurs when a population undergoes a drastic reduction in size for at least one generation due to a catastrophic event like natural disasters (earthquakes, floods, and fires) or disease outbreak.
With fewer individuals surviving the bottleneck, there is a high probability that certain alleles are lost, and others become overrepresented purely due to chance. Thus, it causes a loss of genetic diversity. Again, in a bottlenecked population, the surviving individuals are more likely to be homozygous for specific alleles, as fewer different alleles in the gene pool lead to homozygosity in the gene pool.
As a result of this sudden and severe reduction, the surviving individuals may not represent the original genetic diversity of the population. Over time, as the population recovers, the genetic makeup reflects the limited diversity of the survivors, causing a genetic drift.
If we remove a handful of marbles from a jar filled with different colored marbles, each representing a different allele, then the composition of the remaining marbles changes. It reflects the bottleneck in a population caused due to genetic drift. Here are a few examples of the Bottleneck effect found in nature.
- It is believed that the cheetah population experienced a significant reduction in numbers in the past decade or so, possibly due to changes in climate and habitat and interactions with other large predators. This reduction in population size led to a genetic bottleneck, a period during which the number of individuals in a species is sharply reduced, causing a significant loss of genetic diversity.
- The Northern Elephant Seal population experienced a severe bottleneck in the 19th century due to extensive hunting. The surviving population was reduced to a few dozen individuals, leading to a significant loss of genetic diversity. Although the population has since recovered, the effects of the bottleneck are still evident in their genetic makeup.
2. Founder Effect
It occurs when a small group diverges from a large population and establishes a new group in a different geographical area. The genetic makeup of this new group may not perfectly represent the diversity of the original population, leading to unique genetic characteristics in them. As the new population grows, it will carry the hereditary traits of the founders, leading to a unique genetic profile.
The founder effect has many similarities with the bottleneck effect. However, the founder effect is caused by forming a colony (rather than a catastrophe).
- Pingelap Atoll in Micronesia provides a classic example of the founder effect. A typhoon in the 18th century dramatically reduced the population, and the survivors carried a high frequency of a gene causing color blindness. Today, a significant portion of the Pingelapese population has inherited this genetic trait, showcasing the lasting impact of the founder effect.
- As a subset of the sheep population was brought to the isolated island of Åland in the Baltic Sea, the Finnish landrace sheep (originally from Finland) experienced the founder effect. This founding group represented only a fraction of the genetic diversity of the broader Finnish landrace sheep population. Consequently, the Åland sheep population started with a limited set of genetic variants, which became the foundation for subsequent generations.
Effects of Genetic Drift
Loss of variance.
Genetic drift tends to be more pronounced in smaller populations. As a result, chance events can lead to the loss of specific alleles from the gene pool. This reduction in genetic diversity can limit the adaptive potential of a population, making it more susceptible to environmental changes.
Increased Homozygosity in a Population
Genetic drift increases the frequency of homozygous individuals in a population. Homozygosity is found to occur when an individual possesses two identical alleles at a particular gene locus. As genetic drift reduces the number of alleles, individuals are more likely to inherit identical copies from their ancestors.
Loss of Beneficial Alleles and Fixation of Deleterious Alleles
Genetic drift in small populations can increase the risk of inbreeding, as closely related individuals may contribute disproportionately to the next generation. Inbreeding can cause the expression of recessive deleterious alleles and result in reduced fitness and health. Beneficial alleles might be lost, and deleterious alleles might become fixed purely due to chance factors.
Gene Flow vs. Genetic Drift
Gene flow and genetic drift are ways of changing genetic variation within a population with time. There are similarities and differences between the two.
Gene flow , or migration, refers to the movement of genes or alleles between two different populations of the same species. It occurs when individuals from one population migrate and interbreed with individuals from another population. The movement occurs either physically or through the transfer of gametes (pollen, seeds, or spores).
In contrast, genetic drift is the random change in the allele frequencies of a gene in a population due to chance events. It is more pronounced in smaller groups where random sampling can substantially impact allele frequencies. It is driven by random events, such as a population bottleneck or founder effect, which reduces the size of the population.
While gene flow acts on a larger scale, influencing populations through the movement of individuals, genetic drift operates within populations, especially those of smaller sizes.
- Random Genetic Drift – nature.com
- Genetic Drift – Khanacademy.org
- Genetic Drift and Effective Population Size – Nature.com
- Genetic Drift – Plato.stanford.edu
- What is Genetic Drift? – Study.com
Article was last reviewed on Friday, February 23, 2024
Related articles

Leave a Reply Cancel reply
Your email address will not be published. Required fields are marked *
Save my name, email, and website in this browser for the next time I comment.
Popular Articles
Join our Newsletter
Fill your E-mail Address
Related Worksheets
- Privacy Policy
© 2024 ( Science Facts ). All rights reserved. Reproduction in whole or in part without permission is prohibited.
Genetic drift

Genetic drift n., the process of change in the frequency of an allele (gene variant) in a population over time. Credit: Tsaneda, CC BY 3.0.
Table of Contents
Genetic Drift Definition
What is genetic drift in simple terms? The simple definition of genetic drift ( also referred to as Sewall Wright effect or allelic drift ) is a method of changing the population’s frequency of an allele by chance where some individuals with specific allele reproduce more than the others, this process may result in the loss of beneficial alleles or allele fixation of harmful alleles since the gene frequency changes by chance and not because it is a beneficial allele such as the natural selection theory.
Genetic drift contributes to the natural evolution of species, it may lead to the fixation of new alleles that have been recessively fixed in the previous individuals and the development of a new feature in the population . Genetic drifting occurs in all species; however, it is much more significant in small populations in which the number of individuals has been reduced due to a natural disaster ( bottleneck effect ) or when a number of individuals separate from their population to form a new colony ( founder effect ).
Is genetic drift random? In fact, genetic drift is random as it occurs as a result of pure chance, however, it affects small populations significantly unlike large populations that are not susceptible to change due to chance. For example, if a population of 5 members lost 1 member due to chance without leaving any offspring, the population would lose 1/5 of their genetic pool which represents 20 percent of the population’s genes. Alternatively, if one member is lost from a 50-member population, the lost genes would represent 1/50 of their genetic pool which represents only 2 percent of the overall population’s genes. Therefore, large populations are not greatly affected by the impact of genetic drift.
Unlike natural selection, genetic drift outcomes are unpredictable, it is like flipping a coin without knowing which face you will get, gene drifting outcomes are always due to pure chance.
Genetic Drift Examples

Genetic drift, gene flow, mutations, and natural selection are responsible for the change in the gene pool over time. Example of genetic drift: a population of rabbits with alleles B and b , both alleles are present in equal frequencies p = 0.5 and q = 0.5 if 10 parents reproduce the probability of having an offspring with alleles B or b is 0.5; however, by chance, a slight difference in the offspring allele frequency might occur due to random sampling. As a result, in the next generation, the allele frequencies will change slightly to new frequencies by chance to become p = 0.4 and q = 0.6. in the following generations, the random sampling of alleles continues and the change in allele frequency will be greatly different from the initial 0.5 frequency due to genetic drift. Even though genetic drifting is one of the factors that participate in the evolutionary process of the genetic pool by increasing or decreasing a certain allele’s frequency, however, it does not influence the adaptation of individuals to the environment since the affected genes may be harmful or beneficial genes.
What causes genetic drift?
Even though genetic drift is the result of chance, however, if you need to define genetic drift outcomes you should know how it is affected by many factors such as the number of individuals within the population, where the effect of genetic drift is more predominant in small populations, the second factor is the number of individuals contributing in genetic drifting since some individuals do not produce offspring. Another factor is the occurrence of natural disasters that affects the population size; accordingly, these disasters will increase the significance of random genetic drift among the population as well as the change in natural gene flow patterns. Habitat fragmentation is one of the main factors that influence gene drifting where humans live in nonurban habitats so nonhuman populations are dispersed or even eliminated. As a result, gene flow among these populations is decreased while gene drifting becomes more significant.
Humans can change the effect of gene drifting where they can move individuals to new environments voluntarily or accidentally due to urbanization, urbanization affects the less-mobile species to an extent more than mobile species by increasing the rate of gene flow among these species. The gene flow facilitated by humans is usually known as “Human-facilitated gene flow” it may also introduce new genes into the population allowing for new alleles and mutations. Dispersion of populations due to urbanization may decrease the influence of genetic drift since the bottleneck effect will decrease.
Genetic Drift Consequences
What is the role of Genetic drift in evolution? Genetic results in loss of diversity in the genetic pool where fixation of certain genes may be similar to its initial frequency in the population. Another consequence of genetic drifting is the increase in differentiation among populations where different the population of the same species may have different alleles due to genetic drifting, this occurs if the frequency of one allele is fixed in a population, while its frequency decreases in another population since the frequencies of alleles in the genetic pool proceed in different directions.
Sampling error can cause more changes in small populations’ allele frequencies than the large ones. In small populations, alleles are fixed more rapidly than large populations since the gene pool of large populations tends to be more stable. However, Large populations are affected by genetic drift since they become small as a result of a natural disaster that decreases the population size greatly until the conditions improve to allow the regrowth of the population (bottleneck effect) or when a small population leaves the group to establish a new colony when the population is threatened (founder effect). In order to protect a small population from extinction, genetic diversity should be maintained, consequently, genetic drift should be minimized and mutation should be maintained to facilitate adaptation.
Types of Genetic Drift
The genetic drift effect does not depend on the benefits of an allele, since a harmful allele may be fixed and a beneficial allele may be lost by chance. Regardless of their effect, rare recessive genes can become more common by the effect of genetic drift when the population is exposed to a natural disaster (bottleneck effect) or when a group of individuals separates from a population (founder effect) where the effect of genetic drift appears greatly in small populations. In more detail, we are discussing the bottleneck effect vs founder effect:
Population bottleneck

What is the Bottleneck Effect? The bottleneck effect definition is the decrease in the number of individuals in a population due to a natural disaster, genetic drift bottleneck effect usually influences the genetic distribution among the population, therefore, the effect of genetic drift becomes more significant. Consequently, the genetic variation among this population will decrease as the number of mating individuals will decrease.
When genetic drift rate increases in a population this leads to the loss or the fixation of some alleles, this phenomenon is described in terms of a decrease in the genetic effective size. Even though populations experiencing a bottleneck may reproduce and become larger in size again, however, the genetic variation among these populations declines at a rate that represents the size of the disaster until new individuals are introduced into the population through migration or when new mutations occur. The bottleneck biology strength is affected by its size and duration, these factors are calculated mathematically to determine the influence of the bottleneck on the population’s genetic variation.
Founder effect
The founder effect definition in biology is when a small population may be formed due to the founder effect when a small number of individuals leave their population to start a new colony, these individuals do not necessarily cover the whole genetic set of the population; therefore, gene drifting effect is significant within this small population. An example of the founder effect is most commonly found among species of fungi where the spores disperse and colonize in different environments forming new colonies that may not have the same alleles as the population from which they were originated.
Genetic Drift vs Gene Flow
Gene flow is the flow of alleles from one generation to another by means of migration or dispersion, some populations do not usually experience migration or dispersion while others are more flexible, for example, plants and fungi send their pollens or spores away from their population to colonize in different environments. Even though some populations may seem stable, however, they are not as stable as they seem, such as lions that leave their mothers after development to search for a female that is not related to their population. This flow of genes among populations contributes to the change of the gene pool of every population as well as the introduction of new genes to continue the evolutionary process.
Gene drift is counteracted by gene flow since a population does not usually stay small for a long time to be influenced by genetic drift. However, gene flow can counteract the effect of genetic drift only if the population’s gene flow is sufficient to increase the frequency of alleles lost by gene drifting. Gene flow may occur as a result of passive seed dispersal or active migration, studies have shown that only one migrant per generation can increase the genetic differentiation among the population it can also prevent the genetic drifting effect in decreasing the genetic variation among populations. Note that this rule applies only for ideal populations, while the nonideal populations may require more than one migrant to counteract the genetic drift effect.
Genetic Drift in Evolution
Genetic drift contributes to the increase or decrease of a certain allele in each population; therefore, the effect of genetic drift is canceled over the long run in normal populations, however, the effect of genetic drift cannot be canceled if an allele frequency has reached zero unless a mutation produced this allele again. Genetic drifting is important in evolution since it determines the fate of a mutation, it determines whether it will disappear or becomes fixed in the population after a few generations. For nonideal populations Small in size), genetic drift is important even for the common genes.
Normally, if an allele is fixed in one generation it is most likely to increase in the following generations. However, in terms of genetic drifting, what occurs in one generation does not necessarily occur in the following generations, so if one allele increases in one generation, it may increase or decrease in the next generations.
A subdivided population adaptation is a process consisting of two phases, the first phase is genetic drifting where the loss or fixation of some alleles randomly occurs by chance which in turn helps the population to explore new genes, the second phase is characterized by natural selection of the most beneficial genes that were introduced in phase one, these genes are exported to other populations by migration. The genetic drift theory has a significant role in the evolutionary process of individuals where the balance between mutations and gene drifting creates a state in genetic variation. Since mutations introduce new alleles while gene drifting may eliminate or fix the new alleles.
Try to answer the quiz below to check what you have learned so far about genetic drift.
Choose the best answer.
Send Your Results (Optional)

- Masel, J. (2011). Genetic drift. Current Biology, 21(20), R837-R838.
- Kliman, R. M. (2016). Encyclopedia of evolutionary biology. Academic Press.
- Maloy, S., & Hughes, K. (Eds.). (2013). Brenner’s Encyclopedia of Genetics. Academic Press.
- Clark, M, A., Douglas, M., & Choi, J. (2018). Biology 2e. Openstax
©BiologyOnline. Content provided and moderated by BiologyOnline Editors.
Last updated on June 16th, 2022
You will also like...

The Gene Pool and Population Genetics

Population Growth and Survivorship
Related articles....

A Look Into Natural Selection and its Mechanisms

Encyclopedia of Animal Cognition and Behavior pp 1–2 Cite as
Genetic Drift
- Neelabh 3
- Living reference work entry
- First Online: 26 February 2019
78 Accesses
Random sampling of the organisms in a population leads to changes in the allelic frequencies. This process is termed genetic drift or allelic drift or the Sewall Wright effect (Goodhart 1963 ; Fisher and Ford 1950 ). The source of the change in frequency in case of genetic drift is just “chance.” In other words, unlike natural selection, no environmental factors influence the genetic drift to occur. Genetic drift is one of the basic mechanisms of evolution along with natural selection, migration, and mutation.
Another important feature related to genetic drift is that it is more evident in smaller populations. The reason behind this is that, in small populations, alleles that are present in lower frequencies face a greater chance of being lost than that of higher frequency alleles. Therefore, if the force of the genetic drift is strong enough, it might result in a complete removal of those alleles, leading to a loss in the genetic diversity of the population.
Population bottlenecks and...
This is a preview of subscription content, log in via an institution .
Fisher, R. A., & Ford, E. B. (1950). The “Sewall Wright effect”. Heredity, 4 , 117–119.
Google Scholar
Goodhart, C. B. (1963). The Sewall Wright effect. The American Naturalist, 897 , 407–409.
Goodwin, S. B., Cohen, B. A., & Fry, W. E. (1994). Panglobal distribution of a single clonal lineage of the Irish potato famine fungus. Proceedings of the National Academy of Sciences, 91 (24), 11591–11595.
Hedrick, P. W. (2009). Conservation genetics and North American bison ( Bison bison ). Journal of Heredity, 100 (4), 411–420.
Milgroom, M. G., Wang, K., Zhou, Y., Lipari, S. E., & Kaneko, S. (1996). Intercontinental population structure of the chestnut blight fungus, Cryphonectriaparasitica . Mycologia, 88 , 179–190.
Peltonen, L. (2001). Founder effect. In Encyclopedia of genetics (pp. 724–726). Academic Press, Cambridge, Massachusetts.
Download references
Author information
Authors and affiliations.
Department of Zoology (MMV), Institute of Science, Banaras Hindu University, Varanasi, India
You can also search for this author in PubMed Google Scholar
Corresponding author
Correspondence to Neelabh .
Editor information
Editors and affiliations.
Oakland University , Rochester, MI, USA
Jennifer Vonk
Department of Psychology, Oakland University Department of Psychology, Rochester, MI, USA
Todd Shackelford
Section Editor information
York University, Toronto, ON, Canada
Suzanne MacDonald
Rights and permissions
Reprints and permissions
Copyright information
© 2019 Springer Nature Switzerland AG
About this entry
Cite this entry.
Neelabh (2019). Genetic Drift. In: Vonk, J., Shackelford, T. (eds) Encyclopedia of Animal Cognition and Behavior. Springer, Cham. https://doi.org/10.1007/978-3-319-47829-6_1912-1
Download citation
DOI : https://doi.org/10.1007/978-3-319-47829-6_1912-1
Received : 11 February 2019
Accepted : 13 February 2019
Published : 26 February 2019
Publisher Name : Springer, Cham
Print ISBN : 978-3-319-47829-6
Online ISBN : 978-3-319-47829-6
eBook Packages : Springer Reference Behavioral Science and Psychology Reference Module Humanities and Social Sciences Reference Module Business, Economics and Social Sciences
- Publish with us
Policies and ethics
- Find a journal
- Track your research
Learn Anthropology
No products in the cart.
Username or Email Address
Remember Me Forgot Password?
A link to set a new password will be sent to your email address.
Your personal data will be used to support your experience throughout this website, to manage access to your account, and for other purposes described in our privacy policy .
Get New Password -->
Genetic Drift
- Last Updated: Jul 26, 2023
Genetic drift is a fundamental principle in evolutionary biology. It is a stochastic, non-selective process of allele frequency changes in a population. The process results from random occurrences or chance events, independent of the genotype’s selective advantage or disadvantage. Understanding genetic drift is essential to appreciate the full complexity of evolution and its influence on genetic diversity [1] .

The Mechanism of Genetic Drift
In essence, genetic drift is a process of random sampling. Every generation, alleles are sampled from the existing gene pool to create the next generation. Due to random chance, some alleles get over or underrepresented, leading to changes in their frequency in the population [2] .
Effects on Small Populations
Genetic drift has the most substantial effect on small populations. Small populations are more susceptible to the whims of chance, making it more likely that allele frequencies will deviate from generation to generation. This randomness could potentially lead to the disappearance of some alleles, thereby decreasing genetic diversity.
Bottleneck Effect and Founder Effect
Two special cases of genetic drift often observed are the bottleneck effect and the founder effect:
- Bottleneck Effect : Occurs when a population size significantly decreases due to a catastrophic event or environmental change, leaving a small, random assortment of individuals. The event can lead to genetic drift, where the remaining population has reduced genetic diversity [3] .
- Founder Effect : Occurs when a new population is established by a few individuals. The new population’s gene pool isn’t representative of the source population, leading to genetic drift [4] .
The Implications of Genetic Drift
Genetic diversity.
Genetic drift can significantly reduce genetic diversity, especially in small populations. This is because random fluctuations can inadvertently lead to the loss of certain alleles over time.
Genetic drift is also an important factor in speciation – the process by which new species arise. When a small population is isolated from its parent population (for instance, by geographic barriers), genetic drift can result in significant genetic differences over time, leading to the emergence of a new species [5] .
Genetic Drift vs. Natural Selection
While both genetic drift and natural selection lead to evolutionary changes, they operate differently:
- Genetic drift involves random changes in allele frequencies, regardless of their impact on fitness.
- Natural Selection involves changes in allele frequencies based on the advantage or disadvantage they confer on an organism’s ability to survive and reproduce.
Mitigating the Effects of Genetic Drift
Understanding the principles of genetic drift has allowed us to develop strategies to preserve genetic diversity, especially in endangered species. Conservation efforts often focus on maintaining large population sizes to reduce the impact of genetic drift and sustain the genetic diversity necessary for species’ long-term survival [6] .
Interactions of Genetic Drift with Other Evolutionary Forces
While genetic drift is powerful in its own right, its effects are often influenced by interaction with other evolutionary forces, notably natural selection, gene flow, and mutation.
Genetic Drift and Natural Selection
As mentioned previously, genetic drift and natural selection are different but intertwined mechanisms that can alter the genetic makeup of a population. While natural selection is a deterministic process favouring alleles that increase an individual’s survival and reproductive success, genetic drift is entirely stochastic, with allele frequencies changing purely by chance. The interplay of these two forces can lead to a complex evolutionary landscape.
Genetic Drift and Gene Flow
Gene flow is the exchange of genes between populations. When individuals from different populations mate, they mix their genetic material, introducing new alleles into each other’s populations. This exchange can counteract the effects of genetic drift, maintaining or increasing genetic diversity. However, if gene flow is restricted (for example, by geographical barriers), genetic drift can act more powerfully, potentially leading to speciation [7] .
Genetic Drift and Mutation
Mutation, the random alteration of genetic material, introduces new genetic variants into a population. While genetic drift can remove genetic variation by chance, mutation can restore it. This interplay can maintain a population’s genetic diversity over time [8] .
Genetic Drift: A Key Component of Evolutionary Theory
The concept of genetic drift is vital for our understanding of evolution. It shows us that not all evolutionary change is adaptive. It highlights the role of chance and uncertainty in evolution, providing a counterpoint to the deterministic view offered by natural selection. Additionally, it underscores the importance of population size in determining the fate of genetic variants.
The table below summarizes the effects of genetic drift in conjunction with other evolutionary forces:
Genetic drift is a central tenet of evolutionary biology, playing a critical role in shaping the genetic makeup of populations and potentially leading to new species’ emergence. It operates randomly and independently of natural selection, underscoring the unpredictability and complexity of evolutionary processes. Understanding genetic drift not only enriches our knowledge of evolution but also helps guide strategies for conserving biodiversity.
[1] Gillespie, J. H. (2004). Population Genetics: A Concise Guide . Johns Hopkins University Press.
[2] Hartl, D. L., & Clark, A. G. (2007). Principles of population genetics . Sinauer associates.
[3] Nei, M., Maruyama, T., & Chakraborty, R. (1975). The bottleneck effect and genetic variability in populations. Evolution , 1-10.
[4] Mayr, E. (1942). Systematics and the Origin of Species, from the Viewpoint of a Zoologist . Harvard University Press. https://doi.org/10.1073/pnas.0501939102
[5] Futuyma, D. J. (2013). Evolution . Sinauer Associates.
[6] Frankham, R., Ballou, J. D., & Briscoe, D. A. (2002). Introduction to Conservation Genetics . Cambridge University Press.
[7] Slatkin, M. (1987). Gene flow and the geographic structure of natural populations. Science , 236(4803), 787-792.
[8] Charlesworth, B., & Charlesworth, D. (2010). Elements of Evolutionary Genetics . Roberts and Company Publishers.

Anthroholic helps the world learn Anthropology for Free. We strive to provide comprehensive and high quality content for deep understanding of the discipline.
Newsletter Updates
Enter your email address below and subscribe to our newsletter
I accept the Privacy Policy
Related Posts

Leave a Reply Cancel Reply
You must be logged in to post a comment.
- Introduction to Genomics
- Educational Resources
- Policy Issues in Genomics
- The Human Genome Project
- Funding Opportunities
- Funded Programs & Projects
- Division and Program Directors
- Scientific Program Analysts
- Contact by Research Area
- News & Events
- Research Areas
- Research investigators
- Research Projects
- Clinical Research
- Data Tools & Resources
- Genomics & Medicine
- Family Health History
- For Patients & Families
- For Health Professionals
- Jobs at NHGRI
- Training at NHGRI
- Funding for Research Training
- Professional Development Programs
- NHGRI Culture
- Social Media
- Broadcast Media
- Image Gallery
- Press Resources
- Organization
- NHGRI Director
- Mission & Vision
- Policies & Guidance
- Institute Advisors
- Strategic Vision
- Leadership Initiatives
- Diversity, Equity, and Inclusion
- Partner with NHGRI
- Staff Search

Genetic Drift
Genetic drift is a mechanism of evolution characterized by random fluctuations in the frequency of a particular version of a gene (allele) in a population. Though it primarily affects small, isolated populations, the effects of genetic drift can be strong, sometimes causing traits to become overwhelmingly frequent or to disappear from a population.
Genetic Drift. Genetic drift is the change in frequency of an existing gene variant in the population due to random chance. Genetic drift may cause gene variants to disappear completely and thereby reduce genetic variation. It could also cause initially rare alleles to become much more frequent, and even fixed.
Scientific Director
Division of Intramural Research
- Genetic Drift – Definition and Examples
What is Genetic Drift?
Genetic Drift or genetic sampling error or Sewall Wright effect is nothing but a variation of change in the gene pool of a small population . It occurs strictly by chance and no other specific reasons.
In other words, we can say that Genetic Drift leads to evolution owing to chance events. Although we can find its effect to be the strongest in small populations. Nevertheless, it occurs in all populations of non-infinite sizes too.
As we all know that natural preference or natural selection plays a prominent part in the mechanism of evolution. But at times evolution takes place by chance too. In such cases, Genetic Drift takes place whether it makes the population better-suited for its environment or not.
We can also understand it as the change occurring due to the sampling error in the selection of the alleles. It happens in the upcoming generation from the gene pool of the current generation.
Examples of Genetic Drift

To exemplify the genetic drift, let’s take a hypothetical example of a population of 100 rabbits that live in the woods. All the rabbits in the woods have various colours of their coats.
The colours are black, grey, white, tan, brown and red. In the population of the rabbits, the alleles creating the coat colour are spread equally.
Suppose a disease comes into the population of those rabbits and due to this 98 rabbits die. The two rabbits which are left are brown and black. Now, we can say that there has been a Genetic Drift from 6 alleles to only 2 alleles.
It may be surprising to know, but the fact is that GD keeps on happening all the time in the populations. But we cannot easily see it. Many times mutations take place too which have little effect on the organisms.
However, such mutations may pass on when reproduction occurs amongst the organisms. Also, in some cases when the organism could not survive, the mutation does not pass.
Let us take an example of GD in the human population assuming about a married couple in which the male is having brown eyes and the female is having green eyes.
If the green allele is dominant, even though there is a 50% chance of brown eyes too, the couple might have children with green eyes by chance. GD may take place in birds too.
We can assume a bird to have an allele for two different sizes of the beak. Now it solely depends on the alleles which show up in the offspring thereby causing genetic drift which will result in the disappearance of one of the beak sizes of the bird.
Genetic Drift not only occurs in animals but it takes place in plants too. Let us exemplify this by taking the example of a plant that produces red and yellow flowers.
Now assume that a fire broke out which destroyed the yellow flowers. Hence, the plant is then left with only red flowers and red being the dominant allele, it will result in the plant reproducing only red flowers.
Customize your course in 30 seconds
Which class are you in.

- Animal Cell – Definition, Structure, Functions
- Plant Adaptations – Types and Examples
- Obelia – Structure, Diagram, Life Cycle
- What Is Adaptation Theory? – Types of Adaptation
- Tadpoles – How to Find Tadpoles?
- Desert Animals – Animals that live in Hot and Dry Desert
- Forest Animals – Types of Forest Animals
- Omnivores Animals – Definition and Examples
- Carnivores Animals – Definition, List, and Examples
12 responses to “Carnivores Animals – Definition, List, and Examples”
How is a Panda carnivorous ! Stupid site for stupid people….LMAO
Yeah, your tote right ’cause pandas eat bugs and bamboo so that makes it an omnivore. NOT a carnivore
THIS SITE IS SOOO PLAIN! And Not Smart(ANS)
no they are not lying over 1,200 giant pandas eat fish and fish is meat
hiiiiiiiiiiiiiiiiiiiiiiiii people
this webite is so trash i hate it
to all mt blind lovers post something in sighn launguage
this site isn’t lying over 1,200 giant panda’s eat fish
my homie left 6 comments on this site
Best site. Helped me a lot! Also, Billy is right. Over 1,200 pandas eat fish and they will eat other animals if it is an absolute must. So, “Panda Lover, If you really love pandas, you got to know more about them. Change your name too. People are gonna think you are a pre-school kid who just learned what pandas are. (You might be one, so..)
Also she said LMAO so, my 6 year old daughter says that while typing.
Leave a Reply Cancel reply
Your email address will not be published. Required fields are marked *
Download the App

Home » Science » Biology » Evolution » Difference Between Gene Flow and Genetic Drift
Difference Between Gene Flow and Genetic Drift
Main difference – gene flow vs genetic drift.
Gene flow and genetic drift are two processes that decrease the genetic variation within a population . However, both gene flow and genetic drift have longer term influences on evolution . The main difference between gene flow and genetic drift is that gene flow refers to the transfer of genes between populations whereas genetic drift is the variation of allele frequencies in small populations, allowing the disappearance of alleles from the population . Gene flow allows the combination of gene pools of two populations. However, gene flow allows the origination of new species by the gene transfer. Since genetic drift is a natural process, it is also called the random drift. Genetic drift occurs through founder effect and bottlenecks.
Key Areas Covered
1. What is Gene Flow – Definition, Features, Role 2. What is Genetic Drift – Definition, Features, Role 3. What are the Similarities Between Gene Flow and Genetic Drift – Outline of Common Features 4. What is the Difference Between Gene Flow and Genetic Drift – Comparison of Key Differences

What is Gene Flow
The gene flow refers to the transfer of genes or alleles from one population to another population. The gene flow is also called gene migration. Gene flow into or out of a population affects the allele frequency of the population. The mobility of individuals is the major cause of gene flow from one population to another population. The greater the mobility of individuals, greater the gene flow. Animals are more mobile than plants. Seeds and pollen grains can be transported for great distances with the aid of wind and animals. Gene flow between two populations allows the populations to combine their gene pools with each other. This may reduce the genetic variation between the two populations. Therefore, gene flow reduces the tendency of speciation . This means gene flow repairs the developing differences, which may lead to producing a daughter species from the existing species. Physical barriers such as impassable mountain ranges, vast deserts, oceans, and man-made barriers may obstruct the gene flow.

Figure 1: Gene Flow
Gene flow can occur between species through hybridization or gene transfer as well. Gene transfer refers to the movement of genetic material across species. It includes horizontal gene transfer, reassortment, and antigenic shift. Both bacteria and viruses mainly undergo gene transfer. Horizontal gene transfer is the transfer of genetic material between unicellular organisms and/or multicellular organisms. Reassortment is the recombination of genetic material of different virus species through the chromosomal crossover . In antigenic shift , two or more virus species combine and form a subtype with a mixture of surface antigens from each combined species. The gene flow is shown in figure 1 .
What is Genetic Drift
Genetic drift is the variation of the relative genotypic frequencies in a small population, allowing the disappearance of particular genes due to the death of individuals or incapability to reproduce. Genetic drift is also called random drift since it is a natural process. Genetic drift can occur in two ways: founder effect and bottlenecks. The recurrence of small population sizes causes the founder effect . A severe reduction of the size of a population is called bottlenecks . Since a new population starts from a small number of individuals, alleles or genotypes of the new population become fixed. Therefore, inbreeding coefficient as well as homozygosity of the population increase as a result of allele fixation. The genetic drift can be seen in populations that undergo regular extinction followed by the recolonization. The effective population size (N e ) determines the magnitude of the genetic drift. The N e can also be defined as the number of inbreeding individuals in a population. N e is used to calculate the amount of genetic drift expected in a particular population. The chance of an allele to be fixed in a population depends on N e and the frequency of distribution of that particular allele within the population. If the frequency of a particular allele is low in a population, the chance of that allele disappearing from that population is high. Only the alleles with high frequencies within a population are fixed through genetic drift. This shows that genetic drift is involved in reducing the genetic diversity of the population.

Figure 2: Genetic Drift
However, genetic drift has long term evolutionary consequences. Accumulation of non-adaptive mutations facilitates the population subdivision or speciation. On the other hand, once the allele fixation occurs in different populations independently, the likelihood of mating between different populations of the same species can be reduced. This allows the emergence of new species. The genetic drift in a rabbit population is shown in figure 2 .
Similarities Between Gene Flow and Genetic Drift
- Both gene flow and genetic drift are involved in reducing the genetic diversity within a population.
- However, both gene flow and genetic drift have long term influences on evolution as well through speciation.
Gene Flow: Gene flow refers to the transfer of genes or alleles from one population to another population.
Genetic Drift: Genetic drift refers to the variation of the relative genotypic frequencies in a small population, allowing the disappearance of particular genes due to death of individuals or incapability to reproduce.
Correlation
Gene Flow: Gene flow allows the alleles to move from one population to the other.
Genetic Drift: Genetic drift is the changes in allele frequencies in a small population.
Gene Flow: Gene flow works on more than one populations at once.
Genetic Drift: Genetic drift works on small populations.
Contribution to Speciation
Gene Flow: Gene transfer, which is a process of gene flow between species, allows the origination of new species.
Genetic Drift: The accumulation of non-adaptive mutations and allele fixation facilitates speciation.
Gene Flow: The transportation of pollen for large distances and the mating of Europeans and native Americans, which results in offspring with mixed features are examples of gene flow.
Genetic Drift: The random deaths of green beetles, leaving brown beetles alive is an example of genetic drift.
Gene flow and genetic drift are two events that reduce the genetic diversity of a population. Gene flow is the transfer of genes from one population to the other. Gene transfer is the gene flow between two different species. Gene transfer allows the emergence of a new species. Genetic drift is the variation of allele frequencies of a small population. Genetic drift allows alleles with high frequencies to become prominent within a population. The main difference between gene flow and genetic drift is the effect of each event on the alleles of a population.
1. Rodriguez, Tommy. “Gene Flow.” Darwin was Right. N.p., n.d. Web. Available here. 30 July 2017. 2. “Genetic Drift.” APSnet. N.p., n.d. Web. Available here. 31 July 2017.
Image Courtesy:
1. “ Genetic drift in a population Figure 19 02 02 ” By OpenStax, Rice University – Textbook content produced by OpenStax, Rice University. (CC BY 4.0) via Commons Wikimedia 2. “Gene flow” By Tsaneda – Gene_flow.jpg at wikieducator (CC BY 3.0) via Commons Wikimedia

About the Author: Lakna
Lakna, a graduate in Molecular Biology and Biochemistry, is a Molecular Biologist and has a broad and keen interest in the discovery of nature related things. She has a keen interest in writing articles regarding science.
You May Also Like These
Leave a reply cancel reply.

- Table of Contents
- New in this Archive
- Chronological
- Editorial Information
- About the SEP
- Editorial Board
- How to Cite the SEP
- Special Characters
- Support the SEP
- PDFs for SEP Friends
- Make a Donation
- SEPIA for Libraries
- Entry Contents
Bibliography
Academic tools.
- Friends PDF Preview
- Author and Citation Info
- Back to Top
Genetic Drift
In the 1950s, a lively debate broke out among biologists that continues to this day, over what might seem like the most unlikely of organisms: the land snail, Cepaea nemoralis. Yet, there are in fact some interesting aspects to C. nemoralis . This species of snail is polymorphic; the snail’s shell varies in color (pink, brown, and yellow) as well as the number of visible bands (anywhere from 0–5). But the colors and bands are not equally distributed across populations. In some populations, pink predominates, whereas in others, yellow or brown, and similarly, some banding numbers are more prevalent in some populations than in others. Thus, not only are there variations within populations (it is rare to find a population that is all one color or where all the snails have the same number of bands), but there are variations between populations. What is the explanation for this distribution of forms? Those whose knowledge of evolution familiarized them only with the theory of natural selection might assume, for example, that in the populations where yellow snails were the most prevalent, it was because they were fitter than the other colors—that there was some environmental factor that favored yellow over brown and pink. And that in the populations where brown snails were the most prevalent, there was some difference in the environment that led them to be favored over yellow and pink snails. But is there some other explanation? Perhaps the distributions are in some sense due to chance, perhaps even in a way that can be modeled mathematically. What would that mean, and how would you determine which explanation was correct? The attempt to develop “chancy” explanations that are alternatives (perhaps complementary alternatives) to those due to natural selection is what led biologists to develop models of genetic drift.
Genetic drift (variously called “random drift”, “random genetic drift”, or sometimes just “drift”) has been a source of ongoing controversy within the philosophy of biology and evolutionary biology communities, to the extent that even the question of what drift is has become controversial. There seems to be agreement that drift is a chance (or probabilistic or statistical) element within population genetics (see entry on population genetics ) and within evolutionary biology more generally, and that the term “random” isn’t invoking indeterminism or any technical mathematical meaning, but that’s about where agreement ends. Yet genetic drift models are a staple topic in population genetics textbooks and research, with genetic drift described as one of the main factors of evolution alongside selection, mutation, and migration. Some claim that genetic drift has played a major role in evolution (particularly molecular evolution), while others claim it to be minor. This article will examine these and other controversies.
In order to break through the logjam of competing definitions of drift, this entry begins with a brief history of the concept, before examining various philosophical claims about the proper characterization of drift and whether it can be distinguished from natural selection; the relation of drift to debates over statisticalism; whether drift can be detected empirically and if so, how; and the proper understanding of drift as a model and as a (purported) law.
1. Origins of the Concept of Genetic Drift
2.1 a historically-derived account of drift: the causal process account, 2.2 reactions to and varieties of sampling-based accounts of drift, 2.3 mathematical approaches to drift, 2.4 other accounts of drift, 3. intersection of genetic drift with statisticalist-causalist debates, 4.1 classic studies, 4.2 studies of drift and molecular evolution, 4.3 recent empirical issues concerning drift, 5. drift as models, drift as a law, 6. conclusions, other internet resources, related entries.
Although Charles Darwin invoked “chance” in various ways in the Origin of Species (Beatty 1984), he seems not to have included a concept of drift in his account. He does note in passing that
[v]ariations neither useful nor injurious would not be affected by natural selection, and would be left either a fluctuating element, as perhaps we see in certain polymorphic species, or would ultimately become fixed, owing to the nature of the organism and the nature of the conditions. (Darwin 1872: 63; see similar claims on p. 120 and p. 176)
As the reader will see, this is tantalizingly similar to contemporary conceptions of drift. But Darwin does not develop the idea further; in particular, he does not tell us why the distributions of such variations would be fluctuating over time or how it is that they would ultimately become fixed. The first serious (and mathematical) treatments of drift are usually traced to two of the founders of population genetics, Sewall Wright and R.A. Fisher, although neither claimed to have developed the ideas behind drift (Beatty 1992). Wright (1951) credits John Gulick (1873) with the genesis of the idea whereas Fisher (1922b) first discussed the idea as derived from the work of A.C. and A.L. Hagedoorn (1921), although Wright (1931a) cites the Hagedoorns too. It is unclear who first uses the term “drift” in this context; it appears as early as Wright (1929). So, let us briefly examine Gulick and the Hagedoorns in order to understand the origins of the term “drift”.
Gulick (1873) points out that with natural selection, one can assume that where different forms are found, different external conditions will also be found (with the different forms having adapted over the course of generations to the different external conditions). However, there seem to be cases (e.g., among snails) where the external conditions are very similar, yet the organismic forms are very different. He notes that these species tend to occupy very small areas, even though there is reason to believe it is not because they lack the ability to migrate further. He then postulates a scenario: Suppose some members of a species migrate to a new area where they are free from competition and largely separated from the original population. New variations will arise in the new population, but unless they are “decidedly malformed”, they will persist. The new population will thus come to differentiate itself from the original population (e.g., with new shades of color or with variations of shape), perhaps rapidly if there is a “preexisting tendency to rapid variation”.
Some points to note here that become relevant in later discussions of drift: 1) Drift is described in contrast to natural selection. 2) The variations increasing in the population are those that are neutral, or at least not severely deleterious. (Note that 1 and 2 are also present in the quote from Darwin above). 3) Drift is associated with small populations (although it is not fully clear why). 4) Drift is associated with the founding of a new population in a new area. 5) Changes in the population are the result of movements of organisms and their tendency to produce new variations, both of which are physical processes and not purely mathematical constructs (something that becomes an issue in later debates). 6) The changes described are of organisms in a population.
Hagedoorn and Hagedoorn (1921) similarly point out that some traits of organisms are “trivial”, i.e., “cannot possibly be accounted for as useful”, such as “the shape and arrangement of small hairs on the seeds of some cereals” (p. 107). They likewise maintain that such traits, which can be stable (“pure”, i.e., fixed) within a species, cannot be the product of natural selection; instead, the Hagedoorns assert, they must be “due to some process which accompanies selection” (p. 108). The Hagedoorns then proceed to describe several ways in which variability in a population can be reduced: a new population is founded which lacks some of the variability of the original population; a population is split in half (with the variability in the daughter populations differing from each other and from the original); and “random sampling” where even though the size of the population remains relatively constant from year to year, only a small fraction successfully reproduce. On this last point, they state,
The group of organisms chosen by fate to become the parents of the next generation is usually, but always occasionally, considerably smaller than the number of individuals of their species. (1921: 120)
Thus, the Hagedoorns endorse points 1–5 above, while describing two additional processes besides #4 (the founding of a new population), namely the splitting of a population and the random sampling of parents. They further explain the relevance of #3 (small populations): “the smaller the group, the more limited its potential variability, the sooner it will be pure altogether” (p. 123). And finally, they maintain that drift can produce fixation (“purity”), or the complete loss of variation within a population, even in the absence of selection. Fisher (1922b) reads the Hagedoorns as claiming that “random survival is a more important factor in limiting the variability of species than preferential survival” (p. 321), a claim that he challenged by attempting to show that such a process would be too slow to overcome the rate of mutation (and thus the introduction of new variability—but he seems to say otherwise in 1922a).
An essay published by Wright in 1931 provides what is perhaps one of the earliest explicit characterizations of drift:
It has seemed to me that another factor should be much more important in keeping the system of gene frequencies from settling into equilibrium. This is the effect of random sampling in a breeding population of limited size. The gene frequencies of one generation may be expected to differ a little from those of the preceding merely by chance . In the course of generations this may bring about important changes, although the farther the drift from the theoretical equilibrium, the greater will be the pressure toward return. (Wright 1931b: 205; emphasis added)
The paper from which this quote is taken was meant to be a summary of a longer paper, also published in 1931 (Wright 1986: 88). In the longer paper (1931a), Wright specifies that the random sampling is of gametes . (Gametes are cells that fuse together during fertilization, such as an egg and a sperm). So, even though Wright (1931a) notes that the Hagedoorns had “urged the importance of such random fixation as a factor in evolution”, and states that Fisher (1922b) had analyzed the issue, has he changed the subject to be random sampling of gametes rather than of “parents” (i.e., organisms)? In short, no: Wright (1932 and elsewhere) makes it clear that he considers drift to encompass both random sampling of gametes and random sampling of organisms. In other words, he has expanded the phenomena that the concept of drift is meant to cover from that discussed by Gulick, the Hagedoorns, and Fisher. But Wright’s 1932 paper also emphasized what would become a persistent confusion between drift and inbreeding; both inbreeding and drift are more significant in small populations, so it can become easy to conflate them. But you can have random sampling of parents (say, through a population split) without inbreeding, and inbreeding without random sampling of parents. That alone shows that drift and inbreeding are not the same. So, not all of the expansions of drift were productive ones.
It should be noted that while Wright and Fisher had numerous back-and-forth discussions and disagreements about each other’s claims concerning the role of drift in evolution (Provine 1986, Skipper 2002), they did not seem to disagree about what drift was . Wright (1948) considered the following to be an “acceptable statement” of his view from Fisher and E.B. Ford:
Great evolutionary importance has been attached by Sewall Wright (1931, 1932, 1935, 1940) to the fact that small shifts in the gene-ratios of all segregating factors will occur from generation to generation owing to the errors of random sampling in the process by which the gametes available in any one generation are chosen to constitute the next. Such chance deviations will, of course, be greater the smaller the isolated populations concerned. (Fisher and Ford 1947)
On the other hand, Wright’s later incorporation of fluctuations in mutation rate, fluctuations in migration, and fluctuations in selection (see, e.g., Wright 1949) as types of drift was challenged by Cain and Currey, who asserted that “the worker on actual examples must classify processes according to their biological significance” and that such lumping together would produce confusion and prevent proper analysis of actual situations (Cain & Currey 1963: 59). They thus urged the use of the term “sampling drift”, which Wright adopted in the fourth volume of his 1978 magnum opus, Evolution and the Genetics of Populations .
In short, drift’s founders exhibit a diversity of views about drift, which John Beatty helpfully describes as follows:
drift is a heterogeneous category of evolutionary causes and effects, whose overall significance relative to other modes of evolution (especially evolution by natural selection) has been greatly disputed. (Beatty 1992: 273)
Potential causes invoked in the discussion above include sampling of gametes, sampling of parents, founding of new populations, splitting of populations, each of which is intensified when populations are small, while potential effects mentioned include fluctuations of gene frequencies from one generation to the next, loss of variants from a population, and fixation of a (possibly non-adapted) type in a population. Are these causes and effects all drift? With that sort of confusing heterogeneity, there is little surprise that the concept has drawn philosophical attention. But at least we have our starting point for philosophical discussion (see Beatty 1992 and Plutynski 2007 for additional historical overview).
2. What Is Drift, and Can It Be Distinguished from Natural Selection?
Philosophers have taken a variety of approaches to characterizing drift and distinguishing it from natural selection, including a causal process approach that derives from the history just presented, approaches that are mathematically derived, and other sorts of approaches. These are discussed in turn.
Reflecting on the historical uses of the term “drift”, Beatty states that
what most of the phenomena so designated [as drift] have in common is one or another biological form of random or indiscriminate sampling, and consequent sampling error. (Beatty 1992: 273; see also Plutynski et al. 2016 on the modern synthesis authors’ agreement on this point)
Let’s begin with indiscriminate sampling. Beatty states that parent sampling is
the process of determining which organisms of one generation will be parents of the next, and how many offspring each parent will have (1984: 188; italics in original)
Beatty maintains that this parent sampling can be discriminate, that is, with regard to physical differences, or indiscriminate, that is, without regard to physical differences (1984: 189). Discriminate parent sampling is generally considered natural selection; indiscriminate parent sampling is random drift. Beatty characterizes gamete sampling similarly, as
the process of determining which of the two genetically different types of gametes produced by a heterozygotic parent is actually contributed to each of its offspring (1984: 189; italics in original)
He continues:
This sort of sampling might be indiscriminate in the sense that any physical difference between the two types of gametes produced by a heterozygote might be irrelevant to whether one or the other is actually contributed to any particular offspring. (1984: 189)
And again, the indiscriminate form of sampling is drift while discriminate gamete sampling would be selection.
Several illustrations of indiscriminate sampling are common in the literature, but some are more helpful than others. One is a hypothetical scenario in which two genetically and phenotypically identical twins are walking together; one is struck by lightning whereas the other lives to reproduce. (The example seems to have its origins in Scriven 1959 and Mills and Beatty 1979, although these authors were making a point about fitness, not about drift). This is an unfortunate illustration in part because it is too easy to get caught up in the question of whether the twins are really genetically and physically identical, but, more importantly, it is misleading because in fact, drift requires heritable variation, just as selection does. The lightning example is also problematic because it makes drift seem exceptional and catastrophic, whereas it is generally considered to be pervasive (i.e., occurring in all populations) and not necessarily due to catastrophic or unusual events. Others (e.g., Matthen and Ariew 2002; Walsh, Lewens, and Ariew 2002) use a series of coin tosses as an illustration of drift, but this could be challenged because it encourages binary thinking, instead of allowing for multiple variants with multiple outcomes, and because it is unclear what the “population” of coin flips amounts to.
A better illustration of drift has its origins in Theodosius Dobzhansky’s (1937) discussion of Dubinin and Romaschoff’s (1932) model, which asks us to imagine an urn filled with different colored balls. If the balls are drawn from the urn without respect to color, e.g., by a person drawing balls while blindfolded, then the balls are being indiscriminately sampled (unlike discriminate sampling, where someone deliberately tries to pick balls of a certain color). If a large sample of balls is taken, we expect the frequencies of colored balls in the sample drawn from the urn to be very close to the frequencies in the urn. If only a small sample of colored balls is drawn from the urn, then our sample may very well have different proportions of colored balls than the urn does. Multiple samplings taken over time, which would correspond to multiple generations, would tend to exhibit a pattern of fluctuating frequencies (recall the quote from Darwin above). The illustration thus models the population, its variants with their physical differences, and gives a clear understanding of the possible outcomes. It is also easily extrapolatable to, e.g., colorblind predators (Hodge 1987) and other indiscriminate sampling agents. It does, however, have some limitations; for example, it lacks an analogue for reproduction, since the balls do not produce offspring, multiple or otherwise.
Although the characterization of drift and selection in terms of indiscriminate and discriminate sampling seems straightforward, with Beatty’s explication of indiscriminate sampling an important clarification of what the Hagedoorns, Fisher, and Wright seem to have meant by “random” sampling, Beatty (1984) raises a problem for the conceptual distinction between natural selection and random drift. The problem is as follows: For every population of organisms in a given environment, with a certain distribution of types and associated fitnesses, there is a range of possible outcomes of natural selection, with some more likely than others. It is, of course, more likely that the fitter organisms will have greater reproductive success in the next generation than the less fit, but it is also possible that they will not. (Darwin repeatedly emphasized this chance element of natural selection). So, what do we say about the outcomes where the less fit outreproduce the more fit? Beatty states:
To the extent that those outcomes are less representative of the physical abilities of those organisms to survive and reproduce in the environment in question, any evolutionary change that results will be more a matter of random drift,
concluding that it is
conceptually difficult to distinguish natural selection from random drift, especially where improbable results of natural selection are concerned (Beatty 1984: 196; emphasis in original)
As will be discussed further below, much of the twentieth century was marked by debates among biologists about the relative importance of drift and selection in evolution. Were those debates at least in part the result of conceptual unclarity? Millstein (2002) argues that we need not accept this inadvertent consequence of Beatty’s argument, and that selection can, in fact, be distinguished from drift. In order to do this, three extensions should be made to Beatty’s account. First, similar to Hodge (1987), Millstein suggests that a proper distinction between drift and selection relies on causation, specifically, that drift processes are indiscriminate sampling processes in which any heritable physical differences between entities (organisms, gametes, etc.) are causally irrelevant to differences in reproductive success, whereas natural selection processes are discriminate sampling processes in which any heritable physical differences between entities (organisms, gametes, etc.) are causally relevant to differences in reproductive success. These more precise characterizations of “indiscriminate sampling” and “discriminate sampling” are intended to replace the metaphorical “sampling” talk, retaining the term “sampling” as a useful shorthand only. Second, we should be careful to distinguish the process of drift from the outcomes that drift produces, and the process of selection from the outcomes that selection produces. (Of course, the importance of distinguishing process from outcome is not a novel insight; what is novel here is its application to the problem of distinguishing drift from selection. The distinction has sometimes been rendered as “process vs. product” rather than “process vs. outcome” in the philosophical literature, but given the teleological and other misleading connotations of “product”, the term “outcome” is preferable and “product” should be avoided). Third, we should characterize drift and selection as processes rather than outcomes (as in the first of the three points). If we do these three things, then drift and selection are conceptually distinct and the problem Beatty raises is dissolved; discriminate sampling processes where unlikely outcomes obtain are still selection processes. On this view, it is further acknowledged that it is possible for drift and selection to produce the same outcomes, which helps explain the persistence of biologists’ debates over the relative importance of drift and selection without making them seem trivial (see Millstein 2002 for additional discussion of Beatty’s arguments).
And what are these drift processes? They are the same physical indiscriminate sampling processes that Gulick, the Hagedoorns, Wright, and Fisher (and later, Kimura, who will be discussed further below) sought to characterize: the sampling of gametes in the formation of zygotes, the sampling of parents, the founding of new populations, and the splitting of populations. (Note that this is not intended to be an exhaustive list). The outcomes are likewise those mentioned by drift’s founders: fluctuations of gene frequencies from one generation to the next, loss of variants from a population, and fixation of a type in a population. (Again, this is not an exhaustive list). Each of these outcomes is affected by population size, as any indiscriminate sampling process is; smaller populations undergoing drift tend to experience greater fluctuations in gene frequencies, a faster loss of variants from the population, and faster fixation of types in a population. Thus, the Beatty/Hodge/Millstein account of drift—the Causal Process Account of Drift—is one that is grounded in the historical development of the term and in biological practice, with the philosopher’s role being one of clarification and elaboration. Christopher Stephens (2004), Robert Northcott (2010), and Chris Haufe (2013) also seem to endorse the bare bones view of drift as a sampling process, if not the Causal Process Account of Drift in all of its details.
However, the Causal Process Account of Drift has not gone unchallenged. Robert Brandon (2005) argues that it “does not map well onto the ways biologists differentiate drift from selection” (2005: 156), that selection and drift are the same process (i.e., sampling), and that the large majority of biological cases are not cases of indiscriminate sampling. He maintains that “Drift is any deviation from the expected levels of reproduction due to sampling error” whereas “Selection is differential reproduction that is due to (and in accord with) expected differences in reproductive success” (2005: 168–9). These definitions include both process and outcome. Millstein (2005) responds to each of these challenges and defends her view over his. For the purposes of this essay, it is important to note, as Millstein (2005) acknowledges, that Brandon is certainly correct in his descriptive claim that many biologists incorporate both process and outcome in their definitions of drift (see Millstein, Skipper, and Dietrich (2009) for examples). Indeed, the plurality of definitions of drift offered by contemporary biologists—some process-oriented, some outcome-oriented, some both, and some alternating within the same work—gives rise to the need for philosophical analysis, even if the result, in the end, is to accept that pluralism. The Causal Process Account of Drift is making a prescriptive claim on the grounds of 1) increased clarity, 2) the ability to conceptually differentiate biologically very different phenomena, such as selection in a fluctuating environment from fluctuating gene frequencies due to indiscriminate sampling, which have the same outcomes, while 3) maintaining a grounding in biological practice and (some) biological usage.
Jessica Pfeifer (2005) weighs in on the disagreement between Millstein and Brandon, arguing that it is reasonable to think that the source of probabilities in natural selection are at least partly a result of abstracting from or ignoring certain features of the environment and that, if this view is adopted, it is not conceptually confused to treat selection and drift as causally distinct. On Pfeifer’s view, drift is caused by the distribution of ignored factors, whereas selection for the trait in question is caused by those features that are not ignored.
Peter Gildenhuys (2009) argues that the term “drift” is used to refer to causal influences over a population that have three features: they are non-interactive, non-pervasive, and indiscriminate (NINPICs). Thus, he endorses drift as indiscriminate sampling; the other modifications he makes to the view seem to stem from thinking that the Causal Process Account precludes drift and selection from co-occurring and from thinking the view needs to account for location (e.g., an organism being in the wrong place at the wrong time) as an irrelevant causal factor. In any case, he acknowledges that his account and the Causal Process Account probably agree in practice over what sorts of things should be characterized as constituting drift. Larry Shapiro and Elliott Sober (2007) also endorse the view that selection and drift are distinct processes, but Sober, at least, has backed off this view in a recent paper co-authored with Hayley Clatterbuck and Richard Lewontin (Clatterbuck, Sober, and Lewontin 2013). Like Gildenhuys, they seem to suggest that if selection and drift are distinct processes that they cannot co-occur, and they seem to think that the mere introduction of finite population size introduces drift.
To be clear, however, there can be indiscriminate sampling processes and discriminate sampling processes occurring in the same population, even with respect to the same trait. For example, in a study of over 900 populations, biologist Maxime Lamotte acknowledged that camouflage gave appropriately colored Cepaea nemoralis land snails a selective advantage in their respective environment while simultaneously maintaining that foundings of new populations are “of considerable importance because of the chance variations in the composition of the first colonizers” (Lamotte 1959: 80); the variations he refers to are variations in the colors of the snail colonizers (Millstein 2009). Moreover, since the Causal Process Account requires that variations be heritable, the non-heritable locations of organisms are simply irrelevant for the purposes of deciding on drift vs. selection; for example, the founders of a new population may all hail from the geographic edge of the original population, but they can still be an indiscriminate sample of the whole. Finally, historically, at least (as discussed above), small population size has always been associated with drift, but it was never the main phenomenon to be represented. Thus, the variant sampling accounts of drift should be evaluated in light of these considerations.
Walsh, Lewens, and Ariew (2002) provide a good entrée into understanding a mathematical approach to drift. They begin by acknowledging the historical uses of the term drift, identifying four: 1) a “Sewall Wright Effect”, 2) a “Hagedoorn Effect”, 3) “Indiscriminate Sampling”, and 4) “The Finiteness of Natural Populations” (with some of their characterizations not fully accurate; e.g., with the first, they conflate Sewall Wright’s Shifting Balance Model where drift plays a role with drift itself). They suggest that these phenomena are “disparate” although they acknowledge, citing Beatty (1984), that the first three can be understood in terms of sampling. But the fourth form, they assert, cannot. They describe the fourth phenomenon as follows:
The Hardy-Weinberg Law says that in infinite populations (of diploid organisms) there is no change in gene frequencies when there is no variation in gene fitnesses. But natural populations are finite in size; often they are small. In finite populations there will always be some non-negligible chance that trait frequencies will diverge from expectation. (Walsh, Lewens, and Ariew 2002: 456)
With respect to this understanding of drift as the Hardy-Weinberg Law conjoined with a finite population, Walsh, Lewens, and Ariew maintain that “there is no larger population literally being sampled” (2002: 459). Instead:
…in these cases what happens is that the distribution of fitnesses in the population yields a prediction concerning the way in which a population will change. Drift is manifested as a difference from the outcome predicted by the fitnesses in the population. The law of large numbers tells us that the likelihood of significant divergence from these predictions is an inverse function of the size of the population. The small size of a population increases the chances of error. (Walsh, Lewens, and Ariew 2002: 459)
Walsh, Lewens, and Ariew then state that this is the common feature that the four types of drift they outline all share; thus drift, on their view, is when a series of births, survivals, deaths, and reproductions diverges from the outcome predicted by differences in fitness. Note that, using the terminology introduced in the previous section, this is a purely outcome-oriented (“outcome-only”) definition of drift. Whatever type of process (if any) might have produced the outcome plays no role in the definition. Matthen (2010) defends a similar characterization of drift: “Departures from expected values are what population geneticists call ‘drift’” (Matthen 2010: 3).
However, notice that the fourth drift “phenomenon” that Walsh, Lewens, and Ariew identify isn’t really a phenomenon. Rather, it refers to a model from population genetics, the Hardy-Weinberg model, and predictions based on that model. It is true that as a consequence of the model, there would be no deviation from fitness expectations with an idealized infinite population (if we make some philosophical assumptions about what happens in infinity; Sober (1984) raises questions about these assumptions), but that there would likely be such deviations with a finite population. But this is a purely mathematical approach. The purely mathematical approach doesn’t ask what phenomena the model is supposed to be modeling and why it is that finite populations are of interest. There is also no explanation of why a Hardy-Weinberg model with an assumption of finite population size should count as drift; perhaps this is because of a philosophical tradition of referring to this as drift (e.g., some parts of Sober 1984 read that way), or perhaps it is because of the previously-noted longstanding association between drift and small population sizes. In contrast, on the Causal Process Account of Drift, drift would occur even in infinite populations as long as there is indiscriminate sampling; drift-like outcomes (deviation from fitness expectations) are not required.
Millstein, Skipper, and Dietrich (2009) challenge the outcome-only approach to defining drift and the purely mathematical approach that it is based on. Thus, there seem to be some meta-disagreements between the process-only and outcome-only approaches to drift; should our definitions of drift look to the history of biology or should they look to mathematics? Should we glean our understandings of drift from the phenomena that biologists have sought to understand (the phenomena that they developed models for) or from the models alone? If from the models alone, are we inadvertently drawing on the measure or operationalization of drift rather than drift itself (Plutynski 2007)? Differing answers to these questions seem to be at the heart of the differing definitions of drift.
If one doesn’t interpret the model purely mathematically, but instead considers what phenomenon the Hardy-Weinberg Principle with-finite-populations is seeking to model, then there is indeed a larger population being sampled. The equation models the changes from one generation to the next; the original generation is the population that is being sampled, and the next generation is the “sample”. That sample may or may not be representative of the original population, e.g., a heterozygote Aa of the original generation may or may not have offspring in the proportions 25% AA , 50% Aa , and 25% aa .
The mathematical-oriented outcome-only definition of drift is often associated with the so-called “statisticalist” approach to evolutionary theory, but since it is simply a definition of drift and not a metaphysical thesis it need not be. For example, a non-statisticalist could consistently hold that natural selection is causal (and thus, evolution would not be purely statistical, as it is on the statisticalist interpretation), but that drift occurs when and only when there is a deviation of the actual outcome from fitness expectation. Indeed, Frédéric Bouchard and Alexander Rosenberg (2004) seem to hold such a view. The intersection between issues of drift and the debates over statisticalism will be discussed further below.
Other accounts of drift are not so easily classified. For example, Timothy Shanahan (1992) argues that conceptually, random drift and natural selection are the ends of a continuum. However, to reach this conclusion, Shanahan must reject heritability as a necessary condition for natural selection. As evolutionary biologist John Endler has argued, this has the effect of trivializing natural selection to the claim that “there are differences among different phenotypes” (Endler 1986: 13).
Grant Ramsey (2013) develops a concept he calls “driftability”, which locates drift in individual organisms rather than in a population. Ramsey points out that the possible lives that an organism can lead is a large and heterogeneous set; thus, the actual life that any particular organism leads will probably not be a representative sample (which he seems to equate to an average) of the set. Differences within the set of will lead to different evolutionary outcomes; this intra-organismic heterogeneity within the set of possible lives of an organism is driftability.
Peter Godfrey-Smith (2009) characterizes drift as changes where two parameters, the smoothness of fitness landscapes and dependence of reproductive character on fitness differences, are low. (However, when he needs to explicate why drift has special importance in small populations and why it can be mathematically described in particular ways, he appeals to indiscriminate sampling. Perhaps, then, indiscriminate sampling is at the core of Godfrey-Smith’s view of drift).
Again, these accounts, which seem to deviate significantly from biological usage and practice, raise meta-philosophical issues about how we ought to go about characterizing a scientific concept like “drift”. How far can the definition of a term deviate from scientific usage and practice and still be considered to be a definition of the same thing?
One could also be a pluralist about drift, arguing that there is reason to accept more than one definition of drift, although it is unclear if anyone has actually endorsed this position. Marshall Abrams seems to come the closest, stating:
If random drift is anything, it is not one thing…The term applies to many effects on populations or organisms which are said to be due to “chance,” and to factors which are thought to help to produce such effects. (2007: 673)
Recent debates about random drift are often entangled with debates over the purported purely statistical (non-causal) nature of evolutionary biology, but the issues are separable. There are issues concerning random drift that do not involve questions of statisticalism (as this article seeks to document), and there are issues concerning statisticalism that do not involve random drift (e.g., much of the literature focuses on natural selection rather than random drift). The statisticalist claim, generally traced to a pair of papers by Walsh, Lewens, and Ariew (2002) and Matthen and Ariew (2002), is essentially that evolution is a population-level phenomenon, and that although there are causes at the level of individual organisms (births, deaths, etc.), there are no causes at the population level, only a statistical summary of the individual events. Note that it is widely acknowledged that the models in evolutionary theory are statistical ones, so the distinctive statisticalist claim is that evolutionary biology is purely statistical. There are at least three alternatives to the statisticalist claim; one challenges the claim that evolutionary biology is a population-level phenomenon, arguing that it is constituted by causes at the level of individual organisms (e.g., Bouchard and Rosenberg 2004), a second defends the view that there are population-level causes (e.g., Millstein 2006; Shapiro and Sober 2007), while a third argues for causes at both levels (Pence 2017).
It was already noted above (see section 2.3 ) that the outcome-only definition of drift is often adopted by statisticalists, but not exclusively so; it can also be endorsed by a causalist who, e.g., believes that natural selection is a causal process but that drift is simply deviation from selective expectations. Perhaps surprisingly, it would also be possible for someone to endorse a version of the Causal Process Account of Drift and yet still accept the basic statisticalist premise—if one thought that drift should be understood in terms of indiscriminate sampling, and also thought that indiscriminate sampling should be understood in terms of causes at the level of individual organisms, then the evolutionary changes wrought by drift would just be be the statistical summation of individual level causes. In short, it would be a mistake to infer one’s position on the statisticalist debate from one’s definition of drift, although such slippage is common and the issues are in truth often entangled.
What, then, are the statisticalist issues that random drift is entangled with? The concerns raised by Walsh, Lewens, and Ariew (2002) and Matthen and Ariew (2002) have their origins in claims made by Sober (1984) in his classic The Nature of Selection . Sober characterizes evolutionary theory as a theory of forces, with its zero-force state described by the Hardy-Weinberg equation of population genetics (see the population genetics entry for an explanation of the equation); in such a state, there is no selection, no mutation, no migration, no meiotic drive, random mating, and infinite population size. Thus, the Hardy-Weinberg equation is an idealized model that never obtains in the real world. It is a bit difficult to see where drift fits into the equation, which is no doubt the source of much of the confusion over how to define drift. In his 1984 book, Sober alternatively characterizes drift in terms of random sampling (the process, which occurs, Sober explains, during gamete formation and the founding of new populations) and sampling error (the outcome, i.e., the deviation from fitness expectations). From the point of view of process, at least some types of sampling could be understood to be part of the Mendelian process. The Mendelian process includes the “process wherein organism produce gametes and gametes produce organisms” (Sober 1984: 35), which Sober says is not treated as a force, but rather as the background against which evolutionary forces are described. But Sober does not take this route. Instead, Sober contrasts drift as sampling error (again, the outcome—presumably introduced when one relaxes Hardy-Weinberg assumptions to allow for finite populations) with selection, mutation, and migration; all are forces, Sober asserts, but drift is a different sort of force. It is not “deterministic” and it does not have a definite direction (although it does have a magnitude, determined by the population size). That is, given trait frequencies and fitness values, Sober suggests, selection predicts a specific outcome for the next generation in a specific direction (and is in this sense “deterministic”), whereas drift could yield an increase any of the types present in the population (and is in this sense directionless). Moreover, Sober states, you cannot say how much drift has contributed a change relative to the change introduced by selection; to do so would be as impossible as trying to say, when flipping a fair coin ten times and obtaining six heads, how much of the result was due to the fairness of the coin and how much was due to the fact that it was tossed ten times. He thus concludes that “if drift is an evolutionary force, it is a force of a different color” (1984: 117); he calls it a force, he says, mainly to indicate its causal role.
These metaphysical claims about drift (and selection and other evolutionary processes—but it is just drift that interests us here) set the stage for the statisticalists’ challenge. Matthen and Ariew (2002) challenge the claim that there is a defensible sense in which drift is a force. Aside from the fact that it does not have a predictable or constant direction (as Sober readily acknowledges), they point to a case of two similar populations subject to the same selective pressures, one in which the trait \(T\) becomes fixed and the other in which the alternative trait \(T'\) becomes fixed. What explains the outcome in the two cases? Exactly the same thing, Matthen and Ariew assert, just as exactly the same coin setup explains two heads and four heads in two series of ten coin tosses. Thus, they seem to suggest, there is no additional cause or force, “drift”. Walsh, Lewens, and Ariew (2002) similarly attack Sober’s claim that drift is a force; following Rosenberg (1994), they assert that “the events that are labelled ‘drift’ events, like lightning strikes, etc. are no different in kind from selection events” (Walsh, Lewens, and Ariew 2002: 457)—and thus, drift is not a distinct sort of force. Moreover, they argue, drift is best interpreted statistically as statistical error (see argument in section 2.3 ), and so again, it is not a force.
To be clear, there is a historical component to the statisticalists’ claims; on their view, Darwin’s evolutionary theory was causal, but with the introduction of population genetics, evolution became purely statistical. (It is sometimes unclear whether the statisticalists are making claims about models/theories or whether they are making ontological claims, or whether they think that the latter can be inferred from the former). Hodge (2016; see also Plutynski et al. 2016) challenges this historical claim by arguing that in fact Darwin—although he liked to think that his epistemological and nomological ideals were in descent from Newton’s—never understood natural selection to be a lawful force along with gravitation or inertia. Hodge further argues that biologists today compare and contrast natural selection with artificial selection and with drift in ways directly descending from Darwin’s view of natural selection as a causal process.
A number of other philosophers have sought to challenge the statisticalists’ claim that drift is not a force. For example, Stephens (2004) defends the claim that drift is both a force and a cause. He asserts that if we understand drift properly as a process of indiscriminate sampling (instead of as an outcome, as the statisticalists do), we can see that it is a force, but that in a population of a given size it always has the same force, regardless of whether there is a large or a small deviation from expectation. (It may be that Sober’s inadvertent vagueness in his 1984 book on the process/outcome question contributed to the confusion, since the statisticalists were responding to Sober). He clarifies that, “because drift is a probabilistic cause, the same causal force can have two different outcomes” (Stephens 2004: 557; emphasis in original). Stephens further defends the claim that drift can be understood as a force or a cause by arguing ( contra Matthen and Ariew) that drift does have a direction, namely, eliminating heterozygosity; that it can “make a difference” by affecting the probability of evolutionary change (more in small populations, less in large); and that we can cogently speak of the relative importance of drift in an ensemble of populations or in a population, albeit not at the level of individuals (see Walsh 2007 for a response to Stephens 2004, 2010 and Gildenhuys 2014 for a response to his response).
Brandon (2006), however, argues that “eliminating heterozygosity” is not sufficient to show that drift is directional, given that (as Stephens would readily acknowledge) if there were two alleles at a locus, beginning at equal frequencies, we could not predict which of the two alleles would go to fixation, only that one of them would; Brandon likens this to saying that “a 20-Newton force is acting on object A ”, (2006: 325), which, he seems to imply, is not a directional claim. Moreover, he argues, drift is not a separate process from selection. Thus, drift, on Brandon’s view, is not a force (but it is a law; more on that below). Joshua Filler (2009) responds to Brandon by defending a more elaborated account of what a force is (in part drawing on Bigelow et al. 1988); on that elaborated account, drift’s direction can be seen as less specific than selection, mutation, and migration, but it gives some directional information. Thus, Filler argues, it can be seen as less “forcelike” than selection, mutation, and migration, but it is still a force. Charles Pence (2017), however, worries that Filler’s modifications to our understanding of “force” may be ad hoc , so he offers an alternative defense of the claim that drift is a force. First, Pence argues, we already countenance forces that have stochastically specified directions, such as Brownian motion, and drift is analogous. Second, we can create an evolutionary thought experiment where drift is absent, showing that it is not constitutive of all evolving systems; this is intended to respond to Brandon’s claim that drift is not separable. Importantly, however, Pence points out that there are essential questions that the entire debate has yet to address: “what exactly the use of ‘forces’ is to do for us, and when explanations utilizing a force metaphor are useful or perspicuous” (Pence 2017: 1975).
On the other hand, Kenneth Reisman and Patrick Forber (2005) separate the question of whether drift is a force from the question of whether drift is a cause, arguing only for the latter while not taking a stand on the former. In a related paper, they rely on Woodward’s (2003) manipulability account of causation and a 1957 study of drift by Dobzhansky and Pavlovsky to argue that
decreasing the number of founding members in replicate populations produces an increase in the variability of evolutionary outcomes across those replicate populations. (Forber and Reisman 2007: 617)
In other words, they argue that the conditions of the study can be seen to satisfy the conditions of the manipulability account, showing that drift ought to be understood as a (population-level) cause. Shapiro and Sober (2007) state that they endorse Reisman and Forber’s arguments as well as the view of drift as a process. However, it’s unclear whether Shapiro and Sober (or Reisman and Forber) actually do endorse the view of drift as a process. That is, it’s unclear why population size as a causal factor should count as a process.
This is because it’s a bit unclear if drift is actually what is being manipulated in Dobzhansky and Pavlovsky’s (1957) experiments, or if it is just population size—and population size does not seem to be a process. Perhaps Reisman and Forber have simply shown that population size is a causal factor of evolutionary change in populations undergoing drift. They also seem to be taking Woodward’s account very literally, in that they seem to think that the “variable” that has to be manipulated must be a variable in a mathematical model; otherwise, one could (at least in principle) manipulate the sampling process by manipulating the population rather than simply the population size; Clatterbuck’s (2015) account, discussed in section 5 , gives an example of this. (Population size is a variable in mathematical models of drift, whereas sampling, as discussed above, is implicit). One could also conceivably manipulate the environment so that sampling was discriminate rather than indiscriminate or to change the nature of the indiscriminate sampling.
Pierrick Bourrat (2018) offers an alternative response to Reisman and Forber; he implies that Reisman and Forber (2005) have overlooked Woodward’s criterion of invariance when they invoke the manipulability account in support of drift as a population-level cause. Invariance in an intervention, Bourrat states, “measures the extent to which a relationship between two variables satisfying the manipulation condition remains stable or unchanged as various other changes are made in the background of this relationship” (2018: 162), and, again invoking Woodward, he asserts that causal relationships that are more invariant provide better explanations. But, Bourrat argues, causal explanations in terms of individual-level variables are more invariant under intervention than causal explanations in terms of population-level variables, and thus should be preferred for that reason. In other words, similar to Bouchard and Rosenberg (2004) who are briefly mentioned above, Bourrat defends a causalist position, but at the level of individuals rather than at the level of populations. However, Bourrat does concede that it is reasonable to conceive of drift and natural selection as population-level causes when they depend on indeterministic events, as well as for some cases of frequency-dependent selection. Moreover, Bourrat does not discuss Clatterbuck’s (2015) alternative and arguably superior understanding of the relevant manipulation for drift; had he done so, it is not clear that his claim for the superiority of individual-level causal explanations with respect to drift would follow. Again, though, these are disagreements within the causalist camp.
Other challenges to the statisticalists’ claims about drift as a cause neither endorse the view of drift as a force nor drift as a distinct process. For example, Brandon and Ramsey (2007) see drift and selection as “copossible” outcomes of the same process. But because there is a causal process—albeit only one rather than two—the statisticalists’ claims are not upheld. Abrams (2007) likewise seems loathe to adopt “force” talk about drift, at least in a strong realist sense. He argues that “both selection and drift are aspects of a probability distribution over future frequencies of genotypes or phenotypes in a population” (2007: 667). Yet selection and drift can be distinguished:
Selection is the aspect of the distribution controlled by differences in fitness, while drift is the aspect of such a distribution controlled by population size (apart any from effects of population size on fitness). (2007: 667)
This makes it sound as though Abrams is adopting a view of drift akin to that of Reisman and Forber, with its focus on population size, and indeed Abrams does elaborate on the causal role of population size, but (as noted above) his views on drift are more accurately pluralist ones.
More generally, Pence (2017) suggests one way of categorizing the different causalist approaches (although not all of these have explicitly addressed drift):
At the very least, we need to distinguish between (1) the force interpretation, as discussed here; (2) the causal process approach (elaborated most notably by Millstein 2002, 2006, 2013); (3) the causal mechanism approach, first deployed for natural selection by Barros (2008) and building on the work of Machamer et al. (2000); (4) the manipulationist approach, discussed by Reisman and Forber (2005), Forber and Reisman (2007), and Shapiro and Sober (2007), building on the work of Woodward and Hitchcock (2003); and (5) the counterfactual approach, deployed for natural selection by Glennan (2009) and Huneman (2012) and utilizing a notion of counterfactual causal dependence or “relevance”. (Pence 2017: 1983)
4. Detecting Drift Empirically
Throughout the 20 th and 21 st centuries, biologists have struggled to detect drift empirically. In particular, they have experienced challenges in differentiating cases of drift from cases of selection. The Causal Process Account of Drift in particular can help to make sense of why this is so, as will be discussed below. Drift and selection may be different sorts of causal processes (indiscriminate and discriminate sampling, respectively), but they can produce similar outcomes. Biologists have thus struggled to identify distinctive outcomes for drift and selection, only to find underdetermination re-emerge after biologists modified or added to their assumptions about the relevant processes (for a clear characterization of this, see Dietrich and Skipper 2007). Relatedly, biologists have also disagreed over the relative prevalence of drift and selection (Beatty 1995, 1997); as noted above, these disagreements were there almost from the outset with Fisher’s response to the Hagedoorns.
The disagreements over the prevalence of drift and selection began almost immediately after Wright and Fisher incorporated drift into their evolutionary models (see Provine 1986 for an extended discussion of their disagreements, to which this discussion is very much indebted). Wright (1931a, 1932), drawing on his experience with animal breeding, developed the Shifting Balance Theory (SBT), which consisted of three phases; these have been understood either as empirical claims about actual conditions (Provine 1986) or descriptions of the ideal conditions for evolution (Skipper 2002). Skipper provides a clear description of the SBT as Wright would eventually formulate it (see Hodge 2011 for discussion of early formulations):
In the first phase, random genetic drift causes gene frequencies to change and pull subpopulations semi-isolated within the global population into adaptive valleys because random fluctuations in gene frequencies are almost always maladaptive. In phase two, mass selection will then act within subpopulations and increase their fitness, dragging them from adaptive valleys to adaptive peaks. In the third phase, selection between subpopulations, which Wright called interdemic selection, driven by differential dispersion (migration of organisms from more fit subpopulations to less fit subpopulations) would then enable the global population to be raised to its optimal peak. (Skipper 2002: 345)
Drift, then, on Wright’s view, plays an essential role in the evolutionary process. Fisher, by contrast, thought that mass selection on large populations was the predominant and most effective mode of evolution, leaving very little role for drift given the large population sizes. (This disagreement about the relative role of drift in evolution is one of several things that Wright and Fisher disagreed about; e.g., as is clear from the above, they also disagreed about population structure and effective population size. See Skipper 2002, 2009 and Plutynski 2005 for analyses of contemporary biologists who continue to argue the Wright-Fisher debate).
Another, related issue arose concerning the character of the traits that were said to be subject to random drift. Wright’s early writings, in particular, emphasized the nonadaptive character of the traits undergoing drift and produced by drift (as can be see in Skipper’s characterization of Wright’s SBT above). But a number of potential confusions arise here (Provine 1986; Millstein 2009): First, “nonadaptive” does not necessary mean “maladaptive”; it can also mean “neutral”. Second, “nonadaptive” can refer either to the lack of adaptive differences between organisms or species, or it can refer to the lack of adaptation to the environment (but authors are not always clear about which meaning is intended). So, one’s views on the prevalence of drift can be—and historically often have been—tied to one’s beliefs about the prevalence of adaptive traits (in either sense) in populations. Strictly speaking, however, there is no necessary connection between nonadaptive traits and drift; there can be indiscriminate sampling on organisms that are adapted to their environment and which differ in fitness, and the outcomes of that indiscriminate sampling can pull the population in an adaptive direction or a nonadaptive one. Thus, the proper contrast between selection and drift does not map onto “adaptive” and “nonadaptive”, but rather to “adaptive” and “possibly nonadaptive” (but also “possibly adaptive”).
These theoretical considerations were soon followed by field examinations of drift (there were also laboratory studies, but these were less contentious). Two sets of studies of natural populations are particularly notable. One set, referred to in the introduction to this essay, is composed of the studies of the polymorphic land snail, Cepaea nemoralis ; these studies, and debates over the prevalence of drift, began in the 1930s and became quite heated in “The Great Snail Debate” of the 1950s and 1960s (Millstein 2008, 2009 gives more extensive discussion, from which the following is drawn; the moniker “The Great Snail Debate” is due to Provine 1986). Early researchers, most famously Arthur J. Cain and Philip M. Sheppard (1950, 1954), sought to demonstrate the adaptedness of the color and banding morphs of the snails as well as the sizes of the populations in which they lived. But these were challenging to determine; adaptedness was primarily studied indirectly, by seeking correlations between variants and their backgrounds (presuming camouflage and selection by predator), and population sizes varied considerably. And any correlations found were statistical ones, with lots of “noise”. This left the door open for drift; Maxime Lamotte (1959) argued that by examining the populations as a whole, one found greater variation among the small populations than among the large. This was a distinctive signature for drift, but such outcomes are hard to find and require a very special set of circumstances to obtain (large numbers of populations of varying sizes that are easy to count). However, Lamotte’s study is not the only classic evolutionary study to exploit this unique drift outcome; Cavalli-Sforza’s studies of blood groups in humans did so as well (Richardson 2006). Notably, this was not drift as an alternative to selection (Cain and Sheppard had suggested that selection precluded much of a role for drift), but rather, an argument for a substantial role for drift in addition to a substantial role for selection.
Another important set of studies is of the polymorphic Scarlet Tiger Moth, Panaxia dominula (see Provine 1986 for extended discussion, to which the following is indebted). Fisher and Ford deliberately chose P. dominula because of its small population sizes, figuring that if they could make the case for selection against drift in its worst case, they could make a decisive case against Wright, who they saw as advocating a form of nonadaptive evolution. But rather than trying to make a case for selection, as Cain and Sheppard did, Fisher and Ford (1947) tried to make a case against drift by arguing that the fluctuations of gene frequencies across generations were too large to be accounted for by drift and that the size of the fluctuations did not differ between the small and the large populations, as you would expect if they were undergoing drift. Therefore, they concluded, the populations were undergoing selection. Wright’s (1948) reply challenged both the logic of this conclusion (disproving drift does not prove selection) as well as their characterization of his views, but more significantly, he pointed out that Fisher and Ford had not included data on population sizes for the years in which the gene frequency fluctuations were statistically significant, so, in fact, it was possible that the populations had been undergone constrictions in those years, in which case drift would have been able to produce the observed fluctuations. He also pointed out that fluctuations in selection or migration (which he very problematically suggested could be part of an expanded understanding of drift) could have produced the same outcomes.
In short, the P. dominula studies turned on a slightly different but related proposed unique outcome for drift, that of fluctuations over time rather than variation at a time as for C. nemoralis , in both cases comparing small populations to large (as well as considering the capability of populations to produce large fluctuations), but with P. dominula , drawing definitive conclusions ran into difficulty from lack of data concerning the key variable of population size. There is also the recognition that fluctuating selection can produce the same outcome of fluctuations of gene frequencies across generations as drift can, making it difficult to resolve the empirical case. Finally, note that biologists have continued to study both C. nemoralis and P. dominula , with the result that additional processes have been identified that further complicate the empirical analyses (Millstein 2008; Skipper 2009).
As Dietrich (1994) has documented, early work in molecular evolution was focused almost exclusively on selection, something that changed with the advent of the neutral theory of molecular evolution and the work of Motoo Kimura, Jack King, and Thomas Jukes. Dietrich writes:
The basis of its challenge was Kimura’s proposal that most changes detected at the molecular level were not acted upon by natural selection; they were neutral, and the mechanism of their change was random genetic drift. (1994: 22)
To be clear, however, both selection and drift play an essential role in the neutral theory; on Kimura’s view, selection will quickly eliminate the large number of deleterious mutants and fix the small number of advantageous ones, leaving the remaining mutant alleles as neutral, whereupon they undergo a process of drift (Dietrich 2006). Drift would eventually cause the neutral (and nearly neutral) mutants to either go to fixation or be lost (although they would be polymorphic in the meantime), so that observed molecular differences would be
the outcomes of a random process of mutation, which Kimura understood as produced largely from DNA replication error, processes of directional selection, and processes of random drift produced by gamete sampling. (Dietrich 2006: 670)
Many of the debates about the neutral theory contrasted this approach (the “neutralist” approach) to evolution with a more selectionist (or sometimes, “panselectionist”) approach, causing many to refer to a “neutralist-selectionist controversy”.
The advocates of the neutral theory initially proposed it as an unrealistic simple model only to have subsequent data on the prevalence of neutral alleles convince them of its realism. That led to more explicit testing of the neutral theory and debates over the molecular clock (Dietrich 2006). To focus on the first of these, Dietrich (2006) suggests that although the neutral theory promised to generate numerous quantitative testable predictions—and did—testing it has in fact proved difficult. Is this because of the difficulty in identifying outcomes that are unique to drift? Perhaps so. For example, the neutral theory predicted a certain measure of heterozygosity, but when Francisco Ayala and colleagues failed to observe that predicted outcome in a study of natural populations of Drosophila , Jack King responded by saying that many of the assumptions made by the model could be the source of the discrepancy. In other words, as has been well-discussed in the philosophy of science, there are no “crucial experiments” between two theories because theories are always tested together with their assumptions, any one of which can be given up rather than rejecting the theory. Debates over the so-called “molecular clock” can also be understood in terms of the search for distinctive outcomes for drift. At first, it was thought that only drift could make sense of an apparent constant rate of evolution—that only drift would produce such an outcome—yet this time it is the selectionists who modified their assumptions such that their models would also predict a molecular clock outcome (Dietrich and Skipper 2007).
Subsequently, Tomoko Ohta began to argue for a more significant role for weakly selected mutants (Ohta 1973). Essentially, Ohta’s definition of “nearly neutral” includes mutants that are less neutral than Kimura’s “nearly neutral” mutants, and larger amounts of them as well (Dietrich and Millstein 2008). Later refinements included both slightly deleterious and slightly advantageous mutants. This changes the processes that one would expect to be acting. With the (strictly) neutral theory, only drift would act on the neutral mutants. With the nearly neutral theory, the nearly neutral mutations (whether advantageous or deleterious) would subject to very weak selection (discriminate sampling) and to drift in the form of indiscriminate gamete sampling (Dietrich and Millstein 2008). Because the selection is weak, it would be swamped by the effects of drift, but both processes would still be occurring. Ohta believed that the nearly neutral theory could better account for the results found by Ayala and his colleagues (with its large number of relatively rare alleles), and also, that it could better explain some features of the molecular clock (Dietrich and Millstein 2008). However, it has proven much more difficult to find unique outcomes for the nearly neutral theory than the neutral theory. This makes the nearly neutral theory more difficult to test for and less useful as a null hypothesis as compared to the neutral theory, even as it might account for the available data better.
Hayley Clatterbuck, Elliott Sober, and Richard Lewontin (2013) argue that it doesn’t make sense to talk of drift “dominating” selection or being “stronger than” selection. (Among many such claims, this would challenge Dietrich and Millstein’s (2008) claim, discussed in the previous section, that the best way to understand the nearly neutral theory is as weak selection dominated by drift). Clatterbuck, Sober, and Lewontin take selection and drift to be a population-level process or processes (they don’t take a stand on whether there is a one process or two, but they are clearly “causalists” rather than “statisticalists”), but they raise concerns for the way that biologists have understood the value \(Ns\), which is the effective size of the population multiplied by the selection coefficient. After some initial controversies over how to interpret this value, most biologists eventually came to the view that
selection “dominates” drift when \(Ns\) is much greater than some specified number and that drift “dominates” selection when \(Ns\) is much less than that number,
with proposed numbers including 1/4, 1/2, and 1; in between values are thought to be where the two causes are more or less equal (Clatterbuck, Sober, and Lewontin 2013). Clatterbuck, Sober, and Lewontin point out that on the standard picture, the values of N and s do not predict one gene frequency outcome; rather, they predict a probability distribution of possible gene frequency outcomes. But, they argue, you can change this distribution by changing N or by changing s , with values “chosen so that the first change makes more of a difference, or less, or the same, as you please”; thus, it is “arbitrary to focus on comparisons that give drift the upper hand, or that do the same for selection” (Clatterbuck, Sober, and Lewontin 2013: 538). Here, the suggestion seems to be that N represents the drift cause and that s represents the selection cause—although it’s not clear why drift-as-cause should be equated with population-size-as-a-cause—with the conclusion that the causes are not separable. To say that they are separable would, Clatterbuck, Sober, and Lewontin say, would be akin to asking whether the result of four heads of a fair coin tossed ten times is due to the fairness of the coin or the number of tosses. They also suggest that it is impossible to have a population that is not undergoing drift, since even in an infinite population there could be deviation from selective expectations (again, making it hard to separate the causes). Finally, they argue that it doesn’t make sense to say that with identical setups that producing differing outcomes, that selection dominates in some (when the favored allele increases in frequency) and drift in others (when the favored allele decreases in frequency).
Robert Brandon and Lenore Fleming (2014) point out, however, that Clatterbuck, Sober, and Lewontin’s analysis of a seemingly empirical question is not based on an empirical discovery, but rather, a conceptual analysis, and that as a conceptual analysis, it is not fully consistent; that is, they do not consistently treat drift as a causal process, but sometimes treat it as an outcome (as in the last point described in the previous paragraph). Brandon and Fleming cite several recent empirical studies where biologists seek to provide evidence that drift dominates selection, not relying on the simple \(Ns\) to come to that conclusion, and point out that there are other methods for testing the relative strengths of drift and selection, such as the McDonald-Kreitman (MK) test. In Brandon and Fleming’s view, “drift is not a process. It is, however, a predictable result of a process, namely probabilistic sampling” (2014: 581). It is somewhat unclear, however, how on their view the drift outcome (which outcome?) is “swamping” or “overriding” the selective outcome (presumably by the outcome being the cause of some further, as yet unidentified, outcome). Further elaboration on this point would be useful.
It has also been suggested, most notably by John Gillespie (2000a,b, 2001), that many purported instances of genetic drift are in fact due to “genetic draft.” Genetic draft is a process of linked selection (a hitchhiking process) where it is a matter of chance which of two neutral alleles (in a two-locus model) happens to be linked to a site that undergoes an advantageous mutation, and where the timing of these mutations, followed by a rapid selective “sweep” to fixation, is random. As Skipper (2006) discusses, one of the interesting properties of genetic draft is that it can produce an outcome similar to that which one would expect from genetic drift in small populations, namely, a reduction in genetic variation (thus the similar name). Gillespie argues that draft is less sensitive to population size than drift, which leads him to claim that draft is a more significant cause of evolution than drift (Skipper 2006). Thus, draft introduces yet another relative significance debate (Beatty 1995, 1997) to add to the others discussed in this article: drift vs. selection, Wright’s SBT vs. Fisher’s mass selection, neutralist vs. nearly neutralist vs. selectionist molecular evolution. However, it is worth noting that while draft can produce one of the same outcomes that drift can (reduction in heterozygosity), it would not give rise to fluctuating gene frequencies from one generation to the next. So, not all drifty outcomes can be accounted for by draft.
William Provine (2014) has taken the reduction of drift’s role a step further, calling random genetic drift a “fallacy” and arguing that no such phenomenon exists in nature, period. Provine notes that for both Fisher and Wright (and even Kimura), drift was deeply intertwined with inbreeding, to the extent that one was sometimes confused for the other, which seems warranted—or, at least, that the relationship between the two was never fully clarified (see, e.g., Wright 1931a). Provine defines random drift as “fortuitous extinction of genes” at a genic locus on a chromosome. According to Provine,
Wright believed that random sampling of gametes in Mendelism produced “random genetic drift” at every locus [on every chromosome] in small populations, and also that inbreeding led to rampant “random genetic drift” in small populations (Provine 2014: 54).
Here, Provine treats drift as an outcome, and as that outcome seems physically unlikely or even impossible due to gene linkage, there is, on his view, no drift. Wright did assert that
[j]ust because the direction of drift is accidental, the result is a kaleidoscopic shifting of the average characters of the population through predominant types which practically are never repeated, (1931b: 207; see also Wright 1930: 354)
but that makes drift sound like a cause, not an outcome, and it does not explicitly state that the “shifting” occurs at every locus on every chromosome. But even if he did, Wright could certainly have been mistaken about the effects of drift without being mistaken about the phenomenon itself; notably, Provine does not deny the existence of the random (i.e., indiscriminate) sampling of gametes, although he does object to the seeming reification of the term “gene pool” and the neglect of the relevance of chromosomes, points that are well-taken.
As noted above, the phenomenon of drift is represented in mathematical models of population genetics. The standard mathematical model of drift found in textbooks is the Wright-Fisher model, the core of which is the binomial distribution, and it is the model that philosophers of biology typically appeal to. In the Wright-Fisher model—an idealized model, as all models are—there are assumed to be \(N\) diploid adults in a population, mating randomly, with an allele \(A\) that has a frequency of \(p_0\) and an alternate allele at the same locus. The model further assumes that adults produce an infinite number of gametes having the same allele frequency. \(2N\) gametes are drawn from the “gamete pool” at random to constitute the \(N\) diploid individuals of the next generation. (See Ishida and Rosales 2020 for important clarifications on the historical development of the Wright-Fisher model).
However, as Millstein, Skipper, and Dietrich (2009) point out, the Wright-Fisher model is quite idealized, since, of course,
populations do not reproduce by calling in their local statistician and asking her to pick exactly \(2N\) gametes at random (with replacement) and toss them into the next generation (Gillespie 2004: 49)
—but there are more realistic alternatives. John Gillespie’s model, for example, similarly assumes a diploid population of \(N\) members and a two-allele locus, with frequencies \(p\) and \(q = (1 - p)\); it also similarly assumes random mating. Where it differs from the Wright-Fisher model is that each of the \(2Np A\) gametes constitutes the next generation with a random number of offspring gametes. There is no restriction on the distribution of the numbers of offspring nor on the total number of offspring gametes. Thus, unlike the Wright-Fisher model, Gillespie’s model is not tied to binomial sampling, although sampling more generally is still being modeled. Millstein, Skipper, and Dietrich suggest that the philosophical significance of such alternative models (including others such as the Moran model, the Cannings model, or the coalescent) is that we need to be careful about drawing conclusions about drift from any one particular mathematical model, always keeping in mind Giere’s (1988) point that models are built as representations of specific aspects of physical systems.
Clatterbuck (2015) similarly reminds the philosophical community that it is a mistake to just focus on the Wright-Fisher model, highlighting the Eldon–Wakeley model in particular. She emphasizes that different drift models are not predictively equivalent, so that details of the causal network (represented by the differing assumptions of the different models) underlying a population can be seen to change the outcomes of drift. This challenges statisticalists’ assumptions that drift is purely mathematical. Relatedly, she argues that broadening our conception of drift to include alternative models reveals novel ways of intervening on drift that strengthen the causal argument against its statisticalism. For example, by intervening
on the population in such a way as to increase the probability that individuals have a far greater number of offspring (relative to the population size) than allowed by the Wright–Fisher model
we would increase the probability of a neutral allele increasing in frequency (2015: 3501). This is an improvement over Reisman and Forber’s manipulationist arguments (discussed above), since it is more clearly the drift (the sampling process) that is being manipulated rather than just population size.
But perhaps drift is more than just a model (or a set of models); perhaps it is a scientific law of evolutionary biology. Or so Brandon (2006) argues, dubbing a “Principle of Drift” analogous to the Principle of Inertia in that both are intended to be “zero-force” laws. According to the Principle of Drift,
- “A population at equilibrium will tend to drift from that equilibrium unless acted on by an evolutionary force”
- “A population on evolutionary trajectory t , caused by some net evolutionary force F , will tend to depart from the extrapolated path predicted based on F alone (in either direction or magnitude or both) even if no other evolutionary force intervenes, unless F continues to act” (Brandon 2006: 328).
On this view, drift—and thus change—is the default state of evolutionary systems, challenging Sober’s view (discussed previously), that the Hardy-Weinberg Principle characterizes the zero-force state in evolutionary biology. In defending the Principle of Drift, Brandon challenges those who would argue that there are no laws of biology (e.g., Beatty 1995; see cites within), although Brandon (1990) had already done so by defending a Principle of Selection (see also the related work by McShea and Brandon 2010, in which drift is characterized as a special case of what they dub the “Zero-Force Evolutionary Law”, or ZFEL).
Philosophical discussions of random genetic drift have been lively and fruitful. But as they are still relatively recent, there are many issues yet to be explored or explored fully. Although much energy has been diverted toward debating statisticalism, there are many debates over drift that biologists are engaged in that philosophers could profitably weigh in on, both historical and contemporary. With respect to the last two sections of this article in particular (empirical issues concerning drift and models of drift), we may have only begun to scratch the surface.
- Abrams, M., 2007, “How Do Natural Selection and Random Drift Interact?” Philosophy of Science , 74(5): 666–679.
- Beatty, John, 1984, “Chance and Natural Selection”, Philosophy of Science , 51(2): 183–211.
- –––, 1992, “Random Drift”, in Keywords in Evolutionary Biology , edited by Evelyn Fox Keller and Elisabeth A. Lloyd, Cambridge, MA: Harvard University Press, pp. 273–281.
- –––, 1995, “The Evolutionary Contingency Thesis”, in Concepts, Theories, and Rationality in the Biological Sciences , edited by G. Wolters and J.G. Lennox, Pittsburgh: University of Pittsburgh Press, pp. 45–81.
- –––, 1997, “Why Do Biologists Argue Like They Do?” Philosophy of Science , 64(4):S432–S443.
- Bigelow, John, Brian Ellis, and Robert Pargetter, 1988, “Forces”, Philosophy of Science , 55: 614–630.
- Bouchard, Frédéric, and Alex Rosenberg, 2004, “Fitness, Probability, and the Principles of Natural Selection”, British Journal for the Philosophy of Science , 55: 693–712.
- Bourrat, Pierrick, 2018, “Natural Selection and Drift as Individual-Level Causes of Evolution”, Acta Biotheoretica, 66:159–176.
- Brandon, Robert N., 1990, Adaptation and Environment , Princeton, NJ: Princeton University Press.
- –––, 2005, “The Difference between Selection and Drift: A Reply to Millstein”, Biology & Philosophy , 20(1): 153–170.
- –––, 2006, “The Principle of Drift: Biology’s First Law”, The Journal of Philosophy , 103(7): 319–335.
- Brandon, Robert N., and Lenore Fleming, 2014, “Drift Sometimes Dominates Selection, and Vice Versa: A Reply to Clatterbuck, Sober and Lewontin”, Biology and Philosophy , 29: 577–585.
- Brandon, Robert N., and Grant Ramsey, 2007, “What’s Wrong with the Emergentist Statistical Interpretation of Natural Selection and Random Drift?” in The Cambridge Companion to the Philosophy of Biology , edited by David L. Hull and Michael Ruse, Cambridge: Cambridge University Press, pp. 66–84.
- Cain, Arthur J. and John D. Currey, 1963, “Area Effects in Cepaea ”, Philosophical Transactions of the Royal Society of London B , 246: 1–81.
- Cain, Arthur J. and Philip M. Sheppard, 1950, “Selection in the Polymorphic Land Snail Cepaea Nemoralis ”, Heredity , 4(3): 275–294.
- –––, 1954, “Natural Selection in Cepaea ”, Genetics , 39: 89–116.
- Cain, Joe and Michael Ruse (eds.), 2009, Descended from Darwin: Insights into the History of Evolutionary Studies, 1900–1970 , Philadelphia: American Philosophical Society.
- Clatterbuck, Hayley, 2015, “Drift Beyond Wright–Fisher”, Synthese , 192(11): 3487–3507.
- Clatterbuck, Hayley, Elliot Sober, and Richard C. Lewontin, 2013, “Selection Never Dominates Drift (nor Vice Versa)”, Biology and Philosophy , 20(1): 153–170.
- Darwin, Charles, 1872, The Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life , Sixth ed. London: Murray.
- Dietrich, Michael R., 1994, “The Origins of the Neutral Theory of Molecular Evolution”, Journal of the History of Biology , 27(1): 21–59.
- –––, 2006, “Three Perspectives on Neutrality and Drift in Molecular Evolution”, Philosophy of Science , 73(5): 666–677.
- Dietrich, Michael R. and Roberta L. Millstein, 2008, “The Role of Causal Processes in the Neutral and Nearly Neutral Theories”, Philosophy of Science , 75(5): 548–559.
- Dietrich, Michael R. and Robert A. Skipper Jr., 2007, “Manipulating Underdetermination in Scientific Controversy: The Case of the Molecular Clock”, Perspectives on Science , 15: 295–326.
- Dobzhansky, Theodosius, 1937, Genetics and the Origin of Species , New York: Columbia University Press.
- Dobzhansky, Theodosius and Olga Pavlovsky, 1957, “An Experimental Study of Interaction between Genetic Drift and Natural Selection”, Evolution , 11: 311–319.
- Dubinin, N.P. and D.D. Romaschoff, 1932, “Die Genetische Struktur Der Art Und Ihre Evolution”, Biologichesky Zhurnal , 1: 52–95.
- Endler, John A., 1986, Natural Selection in the Wild , Princeton, NJ: Princeton University Press.
- Filler, Joshua, 2009, “Newtonian Forces and Evolutionary Biology: A Problem and Solution for Extending the Force Interpretation”, Philosophy of Science , 76: 774–783.
- Fisher, Ronald A., 1922a, “Darwinian Evolution of Mutations”, Eugenics Review , 14: 31–34.
- –––, 1922b, “On the Dominance Ratio”, Proceedings of the Royal Society of Edinburgh , 42: 321–341.
- Fisher, Ronald A. and E.B. Ford, 1947, “The Spread of a Gene in Natural Conditions in a Colony of the Moth Panaxia Dominula L”, Heredity , 1: 143–174.
- Forber, Patrick and Kenneth Reisman, 2007, “Can There Be Stochastic Evolutionary Causes?”, Philosophy of Science , 74(5): 616–627. doi:10.1086/525608
- Giere, Ronald N., 1988, Explaining Science: A Cognitive Approach , Chicago: University of Chicago Press.
- Gildenhuys, Peter, 2009, “An Explication of the Causal Dimension of Drift”, British Journal for the Philosophy of Science , 60(3): 521–555. doi:10.1093/bjps/axp019
- –––, 2014, “Arbitrariness and Causation in Classical Population Genetics”, British Journal for the Philosophy of Science , 65: 429–444. doi:10.1093/bjps/axs042
- Gillespie, John H., 2000a, “Genetic Drift in an Infinite Population. The Pseudohitchhiking Model”, Genetics , 155(2): 909–19.
- –––, 2000b, “The Neutral Theory in an Infinite Population”, Gene , 261(1): 11–18.
- –––, 2001, “Is the Population Size of a Species Relevant to Its Evolution?” Evolution , 55(11): 2161–2169.
- –––, 2004, Population Genetics: A Concise Guide , Second ed. Baltimore: Johns Hopkins University Press.
- Godfrey-Smith, Peter, 2009, Darwinian Populations and Natural Selection , Oxford: Oxford University Press.
- Gulick, John T, 1873, “On Diversity of Evolution under One Set of External Conditions”, Journal of the Linnean Society of London , 11: 496–505.
- Hagedoorn, A.L. and A.C. Hagedoorn, 1921, On the Relative Value of the Processes Causing Evolution , The Hague: Nijhoff.
- Haufe, Chris, 2013, “From Necessary Chances to Biological Laws”, The British Journal for the Philosophy of Science , 64(2): 279–295.
- Hodge, M.J.S., 1987, “Natural Selection as a Causal, Empirical, and Probabilistic Theory”, in The Probabilistic Revolution, Volume II , edited by Lorenz Krüger, Gerd Gigerenzer and Mary S. Morgan, Cambridge, MA: MIT Press, pp. 233–270.
- –––, 2011, “Darwinism after Mendelism: The Case of Sewall Wright’s Intellectual Synthesis in His Shifting Balance Theory of Evolution (1931)”, Studies in the History and Philosophy of the Biological and Biomedical Sciences , 42(1): 30–39.
- –––, 2016, “Chance and Chances in Darwin’s Early Theorizing and in Darwinian Theory Today”, in Ramsey and Pence 2016: 41–75.
- Ishida, Yoichi and Alirio Rosales, 2020, “The origins of the stochastic theory of population genetics: The Wright- Fisher model”, Studies in History and Philosophy of Biological and Biomedical Sciences , 79(101226): 1–8.
- Lamotte, Maxime, 1959, “Polymorphism of Natural Populations of Cepaea Nemoralis ”, Cold Spring Harbor Symposia on Quantitative Biology , 24: 65–84.
- Matthen, Mohan, 2010, “What Is Drift? A Response to Millstein, Skipper, and Dietrich”, Philosophy and Theory in Biology , 2:e102.
- Matthen, Mohan and André Ariew, 2002, “Two Ways of Thinking About Fitness and Natural Selection”, The Journal of Philosophy , 99: 55–83.
- McShea, Daniel W. and Robert N. Brandon, 2010, Biology’s First Law: The Tendency for Diversity and Complexity to Increase in Evolutionary Systems , Chicago: University of Chicago Press.
- Mills, Susan K. and John Beatty, 1979, “The Propensity Interpretation of Fitness”, Philosophy of Science , 46(2): 263–286.
- Millstein, Roberta L., 2002, “Are Random Drift and Natural Selection Conceptually Distinct?” Biology & Philosophy , 17(1): 33–53.
- –––, 2005, “Selection Vs. Drift: A Response to Brandon’s Reply”, Biology & Philosophy , 20(1): 171–175.
- –––, 2006, “Natural Selection as a Population-Level Causal Process”, British Journal for the Philosophy of Science , 57(4): 627–653.
- –––, 2008, “Distinguishing Drift and Selection Empirically: ‘The Great Snail Debate’ of the 1950s”, Journal of the History of Biology , 41(2): 339–367.
- –––, 2009, “Concepts of Drift and Selection in ‘the Great Snail Debate’ of the 1950s and Early 1960s”, in Cain and Ruse 2009: 271–298.
- Millstein, Roberta L., Robert A. Skipper Jr., and Michael R. Dietrich, 2009, “(Mis)Interpreting Mathematical Models of Drift: Drift as a Physical Process”, Philosophy and Theory in Biology , 1: 1–13.
- Northcott, Robert, 2010, “Walsh on Causes and Evolution”, Philosophy of Science , 77(3): 457–467.
- Ohta, Tomoko, 1973, “Slightly Deleterious Mutant Substitutions in Evolution”, Nature , 246: 96–98.
- Pence, Charles H., 2017, “Is Genetic Drift a Force?” Synthese , 194: 1967–1988. doi:10.1007/s11229-016-1031-2
- Pfeifer, Jessica, 2005, “Why Selection and Drift Might Be Distinct”, Philosophy of Science , 72(5): 1135–1145.
- Plutynski, Anya, 2005, “Parsimony and the Fisher-Wright Debate”, Biology & Philosophy , 20(4): 697–713.
- –––, 2007, “Drift: A Historical and Conceptual Overview”, Biological Theory , 2(2): 156–167.
- Plutynski, Anya, Kenneth Blake Vernon, Lucas John Matthews, and Daniel Molter, 2016, “Chance in the Modern Synthesis”, in Ramsey and Pence 2016: 76–102.
- Provine, William B., 1986, Sewall Wright and Evolutionary Biology , Chicago: University of Chicago.
- –––, 2014, The “Random Genetic Drift” Fallacy , New York: CreateSpace Independent Publishing Platform.
- Ramsey, Grant, 2013, “Driftability”, Synthese , 190(17): 3909–3928.
- Ramsey, Grant and Charles H. Pence (eds.), 2016, Chance in Evolution , Chicago: University of Chicago Press.
- Reisman, Kenneth and Patrick Forber, 2005, “Manipulation and the Causes of Evolution”, Philosophy of Science , 72(5): 1113–1123. doi:10.1086/508120
- Richardson, Robert C., 2006, “Chance and the Patterns of Drift: A Natural Experiment”, Philosophy of Science , 73: 642–654.
- Rosenberg, Alexander, 1994, Instrumental Biology or the Disunity of Science , Chicago: University of Chicago Press.
- Scriven, Michael, 1959, “Explanation and Prediction in Evolutionary Theory”, Science , 130(3374): 477–482. doi:10.1126/science.130.3374.477
- Shanahan, Timothy, 1992, “Selection, Drift, and the Aims of Evolutionary Theory”, in Trees of Life: Essays in Philosophy of Biology , edited by Paul Griffiths, pp. 131–161. Dordrecht: Kluwer.
- Shapiro, Larry and Elliott Sober, 2007, “Epiphenomenalism—the Do’s and the Don’ts”, in Studies in Causality: Historical and Contemporary , edited by G. Wolters and Peter Machamer Pittsburgh, PA: University of Pittsburgh Press, pp. 235–264.
- Skipper Jr., Robert A., 2002, “The Persistence of the R.A. Fisher-Sewall Wright Controversy”, Biology & Philosophy , 17(3): 341–367.
- –––, 2006, “Stochastic Evolutionary Dynamics: Drift Versus Draft”, Philosophy of Science , 73(5): 655–665.
- –––, 2009, “Revisiting the Fisher-Wright Controversy”, in Cain and Ruse 2009: 299–322.
- Sober, Elliott, 1984, The Nature of Selection , Cambridge, MA: MIT Press.
- Stephens, Christopher, 2004, “Selection, Drift, and the ‘Forces’ of Evolution”, Philosophy of Science , 71(4): 550–570.
- –––, 2010, “Forces and Causes in Evolutionary Theory”, Philosophy of Science , 77(5): 716–727.
- Walsh, Denis M., 2007, “The Pomp of Superfluous Causes: The Interpretation of Evolutionary Theory”, Philosophy of Science , 74: 281–303.
- Walsh, Denis M., Tim Lewens, and André Ariew, 2002, “The Trials of Life: Natural Selection and Random Drift”, Philosophy of Science , 69(3): 452–473. doi:10.1086/342454
- Woodward, James, 2003, Making Things Happen: A Theory of Causal Explanation , Oxford: Oxford University Press.
- Wright, Sewall, 1929, “The Evolution of Dominance”, American Naturalist , 63(689): 556–561.
- –––, 1930, “ The Genetical Theory of Natural Selection : A Review”, The Journal of Heredity , 21: 349–356.
- –––, 1931a, “Evolution in Mendelian Populations”, Genetics , 16: 97–159.
- –––, 1931b, “Statistical Theory of Evolution”, Journal of the American Statistical Association , 26(173, Supplement): 201–208.
- –––, 1932, “The Roles of Mutation, Inbreeding, Crossbreeding and Selection in Evolution”, Proceedings of the Sixth International Congress of Genetics , 1: 356–366.
- –––, 1948, “On the Roles of Directed and Random Changes in Gene Frequency in the Genetics of Populations”, Evolution , 2(4): 279–294.
- –––, 1949, “Population Structure in Evolution”, Proceedings of the American Philosophical Society , 93(6): 471–478.
- –––, 1951, “Fisher and Ford on the Sewall Wright Effect”, American Scientist , 39: 452–458.
- –––, 1978, Evolution and the Genetics of Populations, Volume 4: Variability within and among Natural Populations , Chicago: University of Chicago Press.
- –––, 1986, Evolution: Selected Papers , Chicago: University of Chicago Press.
How to cite this entry . Preview the PDF version of this entry at the Friends of the SEP Society . Look up topics and thinkers related to this entry at the Internet Philosophy Ontology Project (InPhO). Enhanced bibliography for this entry at PhilPapers , with links to its database.
- Arthur J. Cain papers
- A Conversation with Luigi Luca Cavalli-Sforza
- Darwin Online
- R.A. Fisher Digital Archive
- Maxime Lamotte
- Sewall Wright papers
- Perspectives on Molecular Evolution
adaptationism | biology: experiment in | biology: philosophy of | causation: and manipulability | causation: probabilistic | chance: versus randomness | Darwinism | evolution | fitness | gene | genetics | genetics: ecological | genetics: genotype/phenotype distinction | genetics: molecular | genetics: population | heritability | laws of nature | levels of organization in biology | models in science | natural selection | natural selection: units and levels of | probability, interpretations of | replication and reproduction | statistics, philosophy of
Acknowledgments
Thanks to Jon Hodge, Charles Pence, Anya Plutynski, and an anonymous referee for extremely helpful comments on an earlier drift, and also to John Beatty, Michael Dietrich, Jon Hodge, and Robert Skipper for ongoing discussions about the nature of drift.
Copyright © 2021 by Roberta L. Millstein < rlmillstein @ ucdavis . edu >
- Accessibility
Support SEP
Mirror sites.
View this site from another server:
- Info about mirror sites
The Stanford Encyclopedia of Philosophy is copyright © 2023 by The Metaphysics Research Lab , Department of Philosophy, Stanford University
Library of Congress Catalog Data: ISSN 1095-5054
IMAGES
VIDEO
COMMENTS
Larger populations are unlikely to change this quickly as a result of genetic drift. For instance, if we followed a population of 1000 rabbits (instead of 10 ), it's much less likely that the b allele would be lost (and that the B allele would reach 100 % frequency, or fixation) after such a short period of time.If only half of the 1000 -rabbit population survived to reproduce ...
Genetic drift is a change in allele frequency in a population, due to a random selection of certain genes. Oftentimes, mutations within the DNA can have no effect on the fitness of an organism. These changes in genetics can increase or decrease in a population, simply due to chance. Genetic Drift Explained. Although variations of genes (also ...
Evo 101. Genetic drift. Genetic drift is one of the basic mechanisms of evolution. In each generation, some individuals may, just by chance, leave behind a few more descendants (and genes, of course!) than other individuals. The genes and other genetic elements of the next generation will be those of the "lucky" individuals, not necessarily ...
Genetic Drift. Genetic drift, also known as allelic drift or the Wright effect, refers to the random change in the allele frequency of a particular gene in a population. It is one of the basic mechanisms of evolution with natural selection. Unlike the latter, it does not depend on an allele's beneficial or harmful effects.
genetic drift, a change in the gene pool of a small population that takes place strictly by chance. Genetic drift can result in genetic traits being lost from a population or becoming widespread in a population without respect to the survival or reproductive value of the alleles involved. A random statistical effect, genetic drift can occur only in small, isolated populations in which the gene ...
Biology definition: Genetic drift is the drifting of the frequency of an allele relative to that of the other alleles in a population over time as a result of a chance or random event. An example where the effect of genetic drift is magnified is the so-called bottleneck effect. Synonyms: allelic drift; Sewall Wright effect.
4. Genetic drift: (chance effects) random change in the frequency of alleles at a locus. short term genetic drift effects: cause changes in allele frequencies, both in small and large populations (Fig. 23.7 (7th) (Fig. 23.4 6th)). The change in allele frequency due to genetic
Roles in Evolution. Spreads "successful" mutation from one population to another through migration of individuals from place of birth to place of reproduction. Opposes the effects of genetic drift (divergence among populations) because it keeps genes flowing throughout; also opposes local selection. With migration, two populations' allele ...
What is genetic drift? Say you have a population of 5,000 people. That makes 10,000 copies of each gene. Imagine a gene where 3,000 of those copies are of one particular allele or type. In the next generation, there won't necessarily be exactly 3,000 copies again. There may be 3,050 or 2,960 copies instead. Some gametes get randomly picked out of all the possible gametes that could have been ...
Genetic Drift. Random sampling of the organisms in a population leads to changes in the allelic frequencies. This process is termed genetic drift or allelic drift or the Sewall Wright effect (Goodhart 1963; Fisher and Ford 1950 ). The source of the change in frequency in case of genetic drift is just "chance.".
Genetic Drift. Genetic drift is a fundamental principle in evolutionary biology. It is a stochastic, non-selective process of allele frequency changes in a population. The process results from random occurrences or chance events, independent of the genotype's selective advantage or disadvantage. Understanding genetic drift is essential to ...
Genetic drift, also known as random genetic drift, allelic drift or the Wright effect, is the change in the frequency of an existing gene variant in a population due to random chance.. Genetic drift may cause gene variants to disappear completely and thereby reduce genetic variation. It can also cause initially rare alleles to become much more frequent and even fixed.
Genetic drift can be defined as the loss of alleles from a population by chance. It is one of the four elements that influence the evolution of a gene pool, along with mutation, gene flow, and natural selection. Genetic Drift. It is also referred to as the "Sewall Wright effect" based on the theory given by the author in 1969.
Genetic drift, like any other evolutionary force, can only operate as an evolutionary force when there is genetic variability. Genetic drift causes its most dramatic and rapid changes in small populations. The chapter consider some examples of founder and bottleneck effects. Disassortative mating can strongly interact with drift-induced linkage ...
Genetic Drift. O. Honnay, in Brenner's Encyclopedia of Genetics (Second Edition), 2013 Genetic Drift and Evolutionary Theory. Genetic drift is at the core of the shifting-balance theory of evolution coined by Sewall Wright where it is part of a two-phase process of adaptation of a subdivided population. In the first phase, genetic drift causes each subdivision to undergo a random walk in ...
Genetic Drift. Genetic drift is the change in frequency of an existing gene variant in the population due to random chance. Genetic drift may cause gene variants to disappear completely and thereby reduce genetic variation. It could also cause initially rare alleles to become much more frequent, and even fixed. Charles Rotimi, Ph.D.
Summary. This chapter contains sections titled: Basic Evolutionary Properties of Genetic Drift. Founder and Bottleneck Effects. Genetic Drift and Disequilibrium. Genetic Drift, Disequilibrium, and System of Mating. Effective Population Size.
Examples of Genetic Drift. To exemplify the genetic drift, let's take a hypothetical example of a population of 100 rabbits that live in the woods. All the rabbits in the woods have various colours of their coats. The colours are black, grey, white, tan, brown and red. In the population of the rabbits, the alleles creating the coat colour are ...
In short, a general theoretical framework for genetic drift is needed. Its absence has often led to . 7 . speculations of adaptation by natural selection when stochasticity could be a simpler explanation (Cabot et . 8 . al. 1993; Alexandrov et al. 2001; Lynch and Conery 2003; Lynch 2007; Ruan et al. 2021a). A solution may . 9
Genetic Drift: The random deaths of green beetles, leaving brown beetles alive is an example of genetic drift. Conclusion. Gene flow and genetic drift are two events that reduce the genetic diversity of a population. Gene flow is the transfer of genes from one population to the other. Gene transfer is the gene flow between two different species.
Genetic Drift Essay; Genetic Drift Essay. Decent Essays. 695 Words ... 0.5 (Figure 2). Genetic drift drift typically behaves differently in each population, as seen in the small population (Figure 1), while the results of natural ... The fruit fly is an excellent model organism due to its short generation time, large offspring numbers, simply ...
Other definitions of genetic drift, such as the Ornstein-Uhlenbeck model, are about non-specific 27 stochastic noises in long-term evolution, unconnected to genetic variation (Szitenberg et al. 2016).
An essay published by Wright in 1931 provides what is perhaps one of the earliest explicit characterizations of drift: ... In short, drift's founders exhibit a diversity of views about drift, which John Beatty helpfully describes as follows: ... ---, 2014, The "Random Genetic Drift" Fallacy, New York: CreateSpace Independent ...